 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Description of an In Vivo Model for the Assessment of Eosinophil Chemoattractants in the Mouse Mauro M Teixeira^+, Timothy J Williams, Paul G Hellewell
Applied Pharmacology, Imperial College of Medicine, National Heart and Lung
Institute, Dovehouse Street, London SW3 6LY Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97189
Sizes of Files:
Text: 15K
Graphics: Line drawings and photographs (jpg) - 16.3K
Tables (jpg) - 30.3K
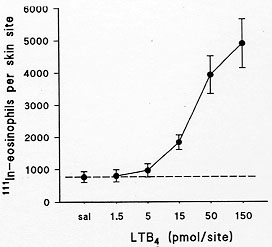
Chemokines (chemoattractant cytokines) induce potent and selective chemotaxis of leukocyte subsets in vitro. Here, we review briefly the chemokines shown to induce eosinophil chemotaxis in vitro and describe a novel model for the study of the ability of chemokines to stimulate eosinophil migration in vivo. Eosinophils were purified from the blood of mice over-expressing the IL-5 gene and labelled with ^111In. Only the C-C chemokines, eotaxin and MIP-1alpha, but not RANTES, MCP-1, MCP-3, MCP-4, MIP-1beta, KC and MIP-2, effectively induced the recruitment of ^111In-eosinophils in mouse skin. We suggest that this mouse model will be useful in assessing the role of endogenously-generated chemokines in mediating eosinophil migration to sites of allergic inflammation in vivo. Key words: chemokines - eosinophils - eotaxin - recruitment Leukocyte migration and chemokines A striking feature of allergic diseases is the presence of an elevated number of eosinophils in tissue without the presence of a significant number of neutrophils (Djukanovic et al. 1990). This preferential accumulation of eosinophils suggests there are specific pathways used by eosinophils, but not by neutrophils, for their selective accumulation in vivo. An understanding of the particular mechanisms modulating eosinophil recruitment in vivo would aid in the development of pharmacological therapies which would block specifically the recruitment of eosinophils, but not that of other leukocytes (Teixeira et al. 1995). Such therapies may be of benefit in allergic diseases where inhibition of eosinophil recruitment is desirable, but without the side-effects of current therapies (eg. steroids) which inhibit leukocyte recruitment indiscriminately (Teixeira et al. 1995) and have other deleterious actions. Recently, it has become clear that a family of newly discovered chemoattractants, namely the chemokines (chemoattractant cytokines), may play an important role in the specific recruitment of leukocytes in vivo. Chemokines are proteins of molecular weight usually ranging from 8 to 10 kDa and amino acid sequence identity of between 20 and 90% (Power & Wells 1996). These proteins have four conserved cysteine residues and, depending on the presence or absence of one amino acid between the first two cysteines adjacent to the N-terminus, are classified in C-C and C-X-C chemokines (Bacon & Schall 1996). More recently, lymphotactin a member of a third sub-family, C chemokines, has been identified which possesses just two cysteine residues (Bacon & Schall 1996). The main function of chemokines appears to be related to the selective activation and recruitment of particular leukocyte subsets, but a number of different roles have been ascribed to these proteins (Bacon & Schall 1996, Oravecz et al. 1996). Eosinophil-active chemokines The action of chemokines on leukocytes is mediated by specific receptors present on the leukocyte surface. Four receptors have now been cloned which mediate the actions of C-X-C chemokines on leukocytes (Mackay 1996). Although C-X-C chemokines preferentially activate neutrophil function and recruitment via stimulation of CXCR1 and CXCR2, more recently receptors for these chemokines have also been described on activated T cells (CXCR3) and the CXCR4 receptor is widely distributed (Mackay 1996). There are five receptors which mediate the actions of C-C chemokines and these receptors are differentially expressed on monocytes, B and T lymphocytes, eosinophils and basophils (Murphy 1996, Power & Wells 1996). For example, eosinophils express high levels of the CCR3 receptor (40,000 to 400,000 receptors per cell) (Ponath et al. 1996a, Daugherty et al. 1996) and also express the CCR1 receptor but only at 1-5% of the levels of CCR3 (Daugherty et al. 1996) It is thus likely that CCR3 is largely responsible for mediating most of the effects of eosinophil-active chemokines (see below, Kita & Gleich 1996). In agreement with their ability to bind to and activate the CCR3 receptor, RANTES, MCP-3, MCP-4 and eotaxin are effective activators of eosinophils in vitro (Kameyoshi et al. 1992, Rot et al. 1992, Jose et al. 1994, Uguccioni et al. 1996). Thus, these chemokines have been shown to elevate intracellular calcium levels in eosinophils and induce chemotaxis, mediator release and the production of oxygen radicals. In addition, it is now evident that elevated levels of these chemokines are found in tissue samples obtained from patients with allergic diseases (Ponath et al. 1996b, GarciaZepeda et al. 1996). Both MCP-2 (Weber et al. 1995) and MIP-1alpha (Rot et al. 1992) have also been shown to activate eosinophils but they are considerably less effective than MCP-3, MCP-4, RANTES and eotaxin (Kita & Gleich, 1996). MCP-2 also appears to recognize a receptor similar to which MCP-3 binds (presumably CCR3) but it has lower affinity (Weber et al. 1995). In human leukocytes, MIP-1alpha does not activate the CCR3 receptor (Ponath et al. 1996b) and is likely to activate eosinophils via the CCR1 receptor. Furthermore, there is evidence to suggest that both RANTES and MCP-3 may also activate the CCR1 receptor in eosinophils inasmuch as RANTES- and MCP-3-induced intracellular calcium elevation in eosinophils is desensitized fully only after prior exposure of these cells to MCP-2 (CCR3) and MIP-1alpha (CCR1) (Weber et al. 1995, Rot et al. 1992). In contrast to the wealth of data demonstrating effects of chemokines in vitro, few animal models exist in which to test comparatively the efficacy and potency of chemokines as eosinophil chemoattractants in vivo. Here, we describe the development of a new model of chemokine-sensitive acute eosinophil recruitment in mouse skin. Eosinophil recruitment in mouse skin Eosinophils were purified from the blood of CBA/Ca mice over-expressing the IL-5 gene. In our transgenic mouse colony, eosinophils account for around 60% of all circulating blood leukocytes (data not shown). Blood was obtained by cardiac puncture (3 to 4 donor mice per experiment) and red blood cells sedimented using Dextran (T500, 1 part of blood to 4 parts of Dextran 1.25%). The leukocyte-rich supernatant was removed, centrifuged (300 g, 7 min) and layered onto a discontinuous 4 layer Percoll (Pharmacia) gradient (densities: 1.070, 1.075, 1.080, 1.085). The gradients were centrifuged at 1,500 g for 25 min at 20 C and the eosinophil/lymphocyte layer collected from the 1.080/1.085 interface. Lymphocytes were removed by using negative immunoselection with rat anti-mouse CD2 and B220 mAbs on a MACS BS column (Miltenyi Biotec LTD). The purified eosinophils (> 95% pure and > 98% viable were then labelled with ^111In and injected intravenously into recipient non-transgenic CBA/Ca mice (1 x 10^6 ^111In-eosinophils/mouse). After 10 min chemoattractants were injected intradermally (i.d.) and ^111In-eosinophil recruitment into skin sites assessed over a period of 2 hr.
Conclusion We have described a new model for the study of eosinophil recruitment in vivo. In this model, eotaxin and MIP-1alpha are effective inducers of eosinophil migration, but eotaxin was significantly more effective than MIP-1alpha. None of the other C-C and C-X-C chemokines tested induced any significant eosinophil recruitment. It is our working hypothesis that chemokines (eg. eotaxin) that activate the CCR3 receptor are fundamental to the process of selective recruitment of eosinophils to tissues in vivo. If that is the case, drugs which block the CCR3 receptor may provide a useful and selective treatment for allergic diseases. We suggest that our mouse model will be useful in assessing the role of endogenously-generated chemokines in mediating eosinophil migration to sites of allergic inflammation in vivo. Moreover, this model should prove useful in the preclinical development of drugs aimed at inhibiting chemokine/CCR3 interactions in vivo. References
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97189b.jpg] [oc97189a.jpg] |
| |||||||||

{kind=link}
{kind=link}