 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
A New Murine Model of Persistent Lung Eosinophilic Inflammation Momtchilo Russo^+, Mario Mariano,^ Sonia Jancar
Departmento de Imunologia, Instituto de Ciências Biomedicas, ICB-III,
Universidade de Sao Paulo, Av. Professor Lineu Prestes 2415, 05508-900 Sao
Paulo, SP, Brasil Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97190
Sizes of Files:
Text: 16.1K
Graphics: Line drawings and photographs (jpg) - 11.5K
Tables (jpg) - 46.2K
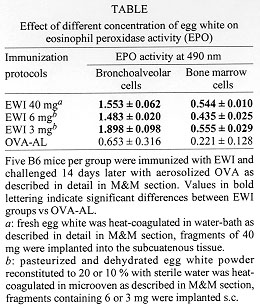
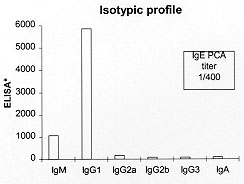
We summarize here the main characteristics of a novel model of pulmonary hypersensitivity. Mice were immunized with a subcutaneous implant of a fragment of heat solidified chicken egg white and 14 days later challenged with ovalbumin given either by aerosol or by intratracheal instillation. This procedure induces a persistent eosinophilic lung inflammation, a marked bone marrow eosinophilia, and Th2-type isotypic profile with histopathological findings that resemble human asthma. Further, this model is simple to perform, reproducible in different strains of mice, does not require adjuvants nor multiple boosters. Based on these characteristics we propose it as a suitable murine model of allergic eosinophilic lung inflammation. Key words: asthma - late-phase reaction - eosinophil - mice - eosinophil peroxidase Asthma is a chronic inflammatory disease of the airways and its incidence is increasing in urban environments. It is characterized by intermittent, reversible airway obstruction and by bronchial hyperreactivity. Nearly a century ago it was observed that eosinophils are the major cell type infiltrating into the airways of asthmatic patients (Ellis 1908). Based on data obtained in animal models of asthma, it has been proposed that T cell-derived cytokines, namely IL-4 and IL-5, play a central role in the induction of lung eosinophilic inflammation. IL-4 is a key cytokine for IgG1 and IgE isotype switching (Drazen et al. 1996), whereas IL-5 is required for the mobilization and migration of eosinophils to the lung (Walker et al. 1991, Nakajima et al. 1992). Indeed, it was shown that IL-4 or IL-5 knockout mice immunized with ovalbumin failed to develop airway hyperresponsive-ness after OVA aerosol challenge (Corry et al. 1996, Foster et al. 1996). Major advances in the understanding of the mechanisms involved in the pathogenesis of asthma came from studies using animal models. A great variety of experimental models have been developed; most of them involve immunization with adjuvants, usually Alum and multiple antigen boosters. We have recently reported that immunization of mice with a subcutaneous implant of fragments of heat-coagulated chicken egg white (EWI) without adjuvant was able to induce a typical late-phase reaction with a prominent eosinophilic infiltrate after ovalbumin challenge into the footpad (Facincone et al. 1997). We have then adapted this model to induce a late-phase reaction in mouse lungs. For this, mice were immunized with EWI implanted into the subcutaneous tissue and challenged with heat aggregated OVA instilled into the trachea 14 days later. It was found that 48 hr after challenge a high percentage of eosinophils was found in the BAL (35%) and increased levels of EPO activity were found in lung homogenates. These results were reproduced in five different mouse strains. Compared to another five models of lung hypersensitivity, the EWI model was the only one to induce eosinophilia which persisted for 30 days. The histopathological findings confirmed the intense eosinophilia and resembled those of human asthma (de Siqueira et al. 1997). In this study we have further characterized this model by analyzing: (a) the efficacy of challenge with aerosolized ovalbumin, (b) the effect of immunization with different concentrations of chicken egg white antigens on airway and bone marrow eosinophilia and (c) the isotypic profile of antibody production. More importantly, we outline the main characteristics of this new experimental model of asthma. MATERIALS AND METHODS Immunizations protocols for the induction of pulmonary eosinophilia - Group of B6 mice were immunized with 100 mg OVA (grade III, Sigma Chemical Co.) adsorbed to 1.4 mg of alum (OVA-Alum) injected intraperitoneally or with a fragment of solidified chicken egg white (EWI) implanted subcutaneously in the dorsal flank of mice through a 5-mm-long cut in the skin. The EWI was prepared in two different ways: (a) separated chicken egg white was placed in a water bath at 100 C for 30 min; the solidified egg white was washed in distilled water, dehydarated in 100% ethanol for 48 hr and cut in to small fragments of 4x2x2 mm weighing around 40 mg each and rehydrated in Dulbecco's PBS for 2 hr before implantation or (b) 10 or 20% solutions of chicken egg white were prepared by dissolving pasteurized and dehydrated chicken egg white (Ito-Avicultura Ind. e Com. S.A., SP, Brazil) in sterile water. A volume of 30 ul of the egg white solutions, containing 3 mg (10%) or 6mg (20%), of egg white, were placed onto glass slides with 12 wells and heat-coagulated in a microwave oven set at maximum potency for 2 min. The solidified fragments of egg white were then fixed in absolute ethanol for 48 hr and rehydrated in Dulbecco's PBS for 2 hr before subcutaneous implantation. Fourteen days after the immunizations the animals were challenged by exposure to an aerosol of OVA (grade III, Sigma) at concentration of 25 mg/ml in 0.9% saline generated by an ultrasonic nebulizer (ICEL US-800, SP, Brazil) delivering particles of 0.5-10 mm of diameter at aproximately 0.75 cc/min for 20 min. Determination of OVA-specific isotypes - Anti-ovalbumin antibodies were assayed by ELISA as previously described (Mengel et al. 1995). Briefly, the Nunc-Immuno Plate MaxiSorp (Inter Med, SP, Brazil) were coated with 2ug of OVA per well, blocked with 0.25% casein in PBS and incubated with two-fold dilutions (1/100 to 1/12,800) of mouse sera. The bound antibodies were revealed by goat anti-mouse IgM, IgG1, IgG2a, IgG2b, IgG3 and IgA followed by peroxidase conjugated rabbit anti-goat-Ig antibody (Southern Biotechnology, Birmingham, AL) and finally by o-orthophe-nylenediamine dihydrochloride (OPD) (Sigma). The reaction was stopped by addition of 50 ul of 4M H2SO4 and the absorbance of the samples determined at 490 nm. ELISA scores were computed by running sums of O.Ds. from 1/100 to 1/12.800 serum dilutions. Each value represents the mean +/- S.D from 5 individual mice. Determination of OVA-specific IgE - OVA-specific IgE antibodies were evaluated by passive cutaneous anaphylaxis (PCA) assay as described by Mota and Wong (1969). Eosinophil peroxidase (EPO) activity - The EPO activity present in bronchoalveolar lavage cells (BAL) and in bone marrow cells was determined with a colorimetric assay as described by Strath et al. (1985) with minor modifications. In brief, cell suspensions were collected as described elsewhere and exposed to Tris-NH4Cl buffer to lyse erytrocytes. The cells were then washed once with PBS and adjusted to 10^6 cells/ml. 100 ml were transferred into wells of 96-well microplate in duplicate. The microplates were then centrifuged at 1,000 rpm at 4 C for 10 min, after which the supernatants were carefully discarded and 100 ml of substrate solution containing 4 mg of OPD in 50mM Tris-HCl, pH 8.0 and 10 ul of H2O2 added to each well. The plates were incubated at room temperature for 15 min. The reaction was stopped by addition of 50 ul of 4M H2SO4 and the absorbance of the samples determined at 490 nm. Bone marrow cell harvesting - Bone marrow cells were obtained by flushing mice femurs with 1 ml of PBS. The cell suspensions were gently homogenized to break up large clumps, submitted to lysing buffer as described above for BAL, counted in a hemacytometer and resuspended to the desired concentration. RESULTS AND DISCUSSION The effect of different concentrations of egg white on pulmonary and bone marrow eosinophilia - The EPO activity assay was employed to detect eosinophils in different compartments (Strath et al. 1985). The levels of EPO activity, a specific enzyme of eosinophils, were determined in bronchoalveolar cells and in bone marrow cells. Groups of five mice were immunized with different concentrations of EWI or with OVA-Alum (for comparison) and the EPO activity was measured seven days after OVA aerosol challenge. As shown in Table, the levels of EPO activity detected in the EWI groups were significantly higher than those obtained in OVA-Alum group. Surprisingly, no significant differences were observed in EPO activity among the groups immunized with EWI at different antigen concentrations. Thus, in these experimental conditions, the intensity of pulmonary and bone marrow eosinophilia was not dependent on the concentration of egg white used for immunization. OVA-specific isotype profile in mice immunized with EWI - The profile of OVA-specific isotype production was evaluated in the serum of five mice immunized with EWI seven days after exposure to 2.5% aerosolized OVA for 20 min. As shown in Fig. this type of immunization induced an increased secretion of IgM, IgG1 and IgE anti-OVA antibodies, but not of IgG2a, IgG2b or IgA isotypes.
The mechanism by which EWI immunization leads to persistent lung eosinophilia remains to be determined. Recently it has been shown that mice that lack the IFN-g receptor present a persistent lung eosinophilic infiltration following OVA challenge (Coyle et al. 1996). We speculate that in the EWI model, the production of IFN-g should be negligible because as shown in Fig., the production of IgG2a or IgG3 isotypes which are known to be dependent on IFN-g production (Stavnezer 1996) was almost absent. Thus it is possible that a selective suppression of IFN-g production may occur in our model. Interestingly, Guery et al. (1996) showed that protein antigens delivered continously by mini-osmotic pump implanted subcutaneously, induced a long-lasting inhibition of IFN-g production. By analogy, the egg white implant may act like a mini pump releasing low amounts of solubilized OVA continously. These possibilities are now under investigation. Finally, based on the data presented here and on data of two recent papers (Facincone et al. 1997, de Siqueira et al. 1997) published by our group, we summarize the most relevant findings in the EWI model. First, it is simple to perform, cheap and reproducible in different strains of mouse and different species, such as rats where it also induces bronchial hyperreactivity (Carvalho et al. in preparation). Second, the model does not require adjuvant or immunization with multiple boosters. Third, it exhibits a Th2-associated isotypic profile and a typical late-phase reaction. Fourth, it induces long lasting pulmonary and bone marrow eosinophilia and lung lesions that may resemble those found in human asthma. ACKNOWLEDGEMENTS To Eliane Gomes for expert technical assistance. This work was supported by grants from FAPESP and CNPq. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97190b.jpg] [oc97190a.jpg] |
| |||||||||

{kind=link}
{kind=link}