 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mechanisms of Cell Accumulation Induced by Mycobacterium bovis BCG Octavio Menezes-de-Lima-Junior, Maria das Gracas MO Henriques^+
Laboratorio de Farmacologia Aplicada, Far-Manguinhos, Fiocruz, Rua
Sizenando Nabuco 100, 21041 250 Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97193
Sizes of Files:
Text: 14.3K
Graphics: Line drawings and photographs (jpg) - 137.4K
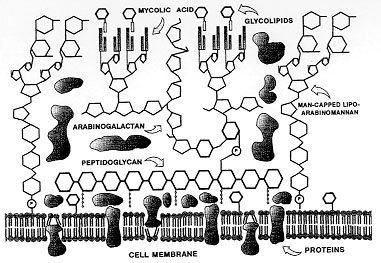
Mycobacteria, specially Mycobacterium tuberculosis are among the micro-organisms that are increasing dramatically the number of infections with death, all over the world. A great number of animal experimental models have been proposed to investigate the mechanisms involved in the host response against these intracellular parasites. Studies of airway infection in guinea-pigs and rabbits, as well as, in mice intravenously infected with BCG have made an important contribution to our understanding of the virulence, pathogenesis and the immunology of mycobacterial infections. Although, there are few models to study the mechanisms of the initial inflammatory process induced by the first contact with the Mycobacteria, and the relevance of the acute generation of inflammatory mediators, cytokines and leukocyte infiltration to the development of the mycobacterial infection. In this work we reviewed our results obtained with a model of M. bovis BCG-induced pleurisy in mice, describing the mechanisms involved in the leukocyte influx induced by BCG at 24 hr. Different mechanisms appear to be related with the influx of neutrophils, eosinophils and mononuclear cells and distinct inflammatory mediators, cytokines and adhesion molecules are involved in the BCG-induced cell accumulation. Keywords: animal models - Mycobacteria - Mycobacterium bovis BCG - inflammation - leukocyte accumulation - pleurisy Over a century ago, Robert Koch identified Mycobacterium tuberculosis as the causative agent of tuberculosis in humans. He was the first to realise that the efficacy of his early therapies with soluble extracts of M. tuberculosis depended largely on the patient's immune-cell response. Mycobacteria, specially M. tuberculosis are among the micro-organisms that are increasing dramatically the number of infections with death, all over the world (Chretien 1995). Mycobacteria are small, aerobic, gram-positive, acid-fast, nonmotile and nonspore-forming bacilli. They consist of a large group including pathogenic, non-pathogenic, and saprophytic species that commonly are of environmental origin (Hines II 1995). Characteristically, mycobacteria presents a complex cell wall associated to the membrane. The cell wall structure and composition (Fig. 1) is related to the ability of the intracellular parasitic species of mycobacteria to survive in the hostile environment of the phagossome of macrophages, as well as with the cytotoxic and immunological effects of these micro-organisms. (Fenton & Vermeulen 1996).
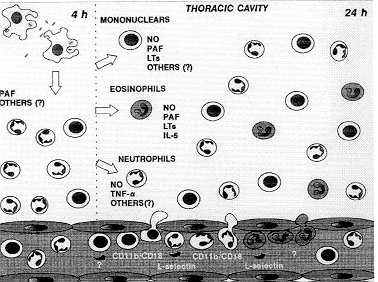
While tuberculosis represents nowadays the most threatening disease caused by intracellular bacteria, our understanding of the cellular and molecular interactions between mycobacteria and host cells is far from complete. The initial events during a primary pulmonary infection with MTC are poorly understanding and there are few models to evaluate the sequence of events that follows the first contact of the host with the mycobacteria. In this article, we review the current state of understanding of the early events that happen during the primary contact with mycobacteria. Animal models Studies with laboratory animals (guinea-pigs and rabbits) have significantly enhanced our understanding of the aetiology and pathogenesis of tuberculosis. Humans, cattle, deer, guinea-pigs and rabbits have similar pathology, but differ in their susceptibility to tuberculosis (Smith & Wiegeshaus 1989). Although studies in guinea-pig and rabbit have made an important contribution to our understanding of the virulence and pathogenesis of tuberculosis, they have limited use for the study of the protective immune response (Dannemberg 1991, Griffin et al. 1995). Since 1970, mice have largely replaced guinea-pigs and rabbits as the animal model to study the immunology of mycobacterial infections. Most of the studies on the role of macrophages and monocytes and its interaction with T cells were done in mice intravenously infected with BCG (Kaufmann 1993). The early events The initial events during a primary pulmonary infection with MTC are poorly understood and there are few models to evaluate the sequence of events that follow the first contact of the host with the mycobacteria. We have recently described the experimental model of mouse pleurisy induced by M. bovis BCG. We used the pleural cavity of the mouse because is a straightforward and well-established model (Henriques et al. 1990, 1996, Bozza et al. 1994). The injection of M. bovis BCG into mouse pleural cavity induces an intense biphasic inflammatory reaction that peaks at 24 hr and 15 days. At 4 hr occurs an influx of neutrophils that is maximal at 24 hr. At this time it is also observed an intense influx of eosinophils and mononuclear cells. Another leukocyte influx is observed at 15 days composed by mononuclear cells and some neutrophils (Menezes-de-Lima-Junior et al. 1997). Neutrophils are the predominant leukocytes to arrive at sites of acute inflammation, capable of defending the host against bacterial infections. Neutrophils accumulation is also intimately associated with oedema formation and the recruitment of other phagocytic leukocytes such as monocytes (Nourshargh 1993). The neutrophil influx induced by i.t. BCG is in accordance to the description of neutrophil migration to peritoneal cavity of rabbits inoculated with BCG (Appelberg 1992) and to rabbit pleural cavity after BCG instillation (Antony et al. 1985). Furthermore, neutrophils are described in pleural exudate of tuberculosis patients (Montgomery & Lemon 1933). Eosinophils are also capable of phagocyte and killing bacteria in vitro, and have a specialised role in the inflammatory process involved in protective responses against helmintic parasites (Nourshargh 1993). Furthermore has been recently reported that eosinophils accumulate in pleural cavity after injection of endotoxin (Bozza et al. 1993) suggesting an involvement of these cells in pulmonary inflammation induced by bacterial products. On the other hand, eosinophil accumulation induced by mycobacteria is poorly described. Although eosinophilia is observed sometimes in broncoalveolar lavage of tuberculosis patients (Vijayan et al. 1992, Nakamura et al. 1993), eosinophil accumulation induced by M. tuberculosis or M. bovis BCG in animal experimental models has not been described so far. However, Castro et al. (1991) have described that M. avium or M. smegmatis are able to induce eosinophil accumulation in mouse air pouch. We have analysed the inflammatory mediators and cytokine involved in eosinophil accumulation induced by BCG comparing with those involved in neutrophil accumulation (Fig. 3). Lipoxygenase products, PAF-acether and interleukin-5 seem to be involved in eosinophil accumulation (Menezes-de Lima-Junior et al. 1996, 1997).
Figure 3: inflammatory mediators, cytokines and adhesion molecules involved in the leukocyte accumulation induced by BCG in mouse pleural cavity.
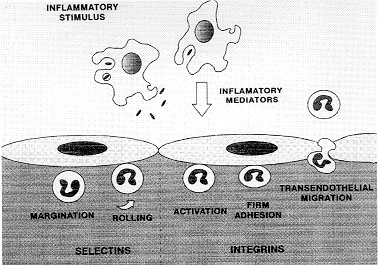
At 24 hr, only dexamethasone or L-NAME were able to inhibit the neutrophil accumulation as well as eosinophil influx (Menezes-de-Lima Junior et al. 1997). The participation of NO in leukocyte chemotaxis in vitro has been investigated (Kaplan et al. 1989, Belenky et al. 1993) although a few in vivo studies have been performed with eosinophil (Teixeira et al. 1993, Ferreira et al. 1996). The sources of NO are probably the macrophages from the pleural cavity. It is known that mouse pleural macrophages can phagocyte M. bovis BCG in vitro (Zlotnik & Crowde 1982). It was also described that peritoneal macrophages from M. bovis BCG-infected mice produces nitric oxide (Saito & Nakano 1996). We have also demonstrated that pleural macrophages from mice injected i.t. with BCG, produces significant levels of NO in contrast to resident macrophages (Werneck-Barroso et al. 1996) The treatment with a neutralising monoclonal antibody (mAb) anti-TNF-a has indicated that this cytokine is involved in neutrophil but not eosinophil migration induced by BCG pleurisy (Menezes-de-Lima-Junior et al. 1997). A number of cytokines can regulate the accumulation and activation of neutrophils and eosinophils. TNF acts on endothelial cells to enhance their interaction with neutrophils and eosinophils. The dose-response relationship and kinetics of TNF-stimulated endothelial cell adhesiveness for neutrophils is similar to that for eosinophils (Nourshargh 1993). However, in our results with BCG induced-pleurisy, TNF-a seems to have a more important effect on neutrophil migration (Menezes-de-Lima-Junior et al. 1997). It was recently demonstrated that TNF-a produced by macrophages in response to PPD (the soluble antigen released from M. tuberculosis) can regulate NO production by these cells (Saito & Nakano 1996). This regulation between NO and TNF-a can be a putative mechanism that is modulating the neutrophil migration induced by BCG in mouse pleural cavity. Involvement of adhesion molecules An important early event in the recruitment of leukocytes from the microcirculation to tissues is their interaction with vascular endothelial cells. In vitro and in vivo studies have suggested that this process involves a sequence of discrete events involving different families of cell adhesion molecules (Springer 1994). In the initial phase, leukocytes marginates to the wall of postcapillary venules and roll along the endothelial cells and this process is mediated by the selectin family of adhesion molecules (Ley et al. 1995). Then, leukocytes must firmly adhere to the vessel wall to migrate to the site of tissue irritation, a step mediated by another family of adhesion molecules the integrins (Fig. 2). The role of adhesion molecules during mycobacterial infection is not clear. We have investigated the role of integrins and L-selectin in acute cell migration induced by i.t. BCG (Fig. 3). The leukocyte integrin, CD11b/CD18 complex seems to have a role in neutrophil and mononuclear cell accumulation. Moreover the integrins CD11b/CD18 and VLA-4, seemed to not be related with eosinophils migration whereas L-selectin appears to be responsible for the neutrophil and eosinophil migration induced by BCG (unpublished results). The role of CD11b/CD18 and L-selectin in neutrophil migration is well demonstrated in vivo with other inflammatory agents (Rossi & Hellewell 1994). However, the adhesion receptors involved in eosinophil recruitment at inflammation in vivo are less clearly understood. Blocking either CD18 or ICAM-1 has been shown to reduce eosinophil accumulation in vivo (Wegner et al. 1990, Teixeira et al. 1994). In addition, blocking VLA-4 in vivo has been shown to reduce eosinophil accumulation in some but not all models (Teixeira et al. 1995). Our results showed that neither the mAb anti- CD11b/CD18 nor the anti-VLA-4 were able to inhibit eosinophil accumulation induced by BCG. One possibility is that both integrins should be simultaneously blocked to inhibit eosinophil migration as it is observed in lung eosinophilia induced by Sephadex (Das et al. 1995). Another possibility is the participation of a1b2 or a4b7 integrin, both described to be expressed in eosinophil surface (Teixeira et al. 1995, Lobb et al. 1996). A recent study in a xenogeneic mesentery model has demonstrated that the rolling of human eosinophil in IL-1b-treated rabbit mesenteric venules is reduced in approximately 40% by anti-L-selectin mAbs (Sriramarao et al. 1994). However a role for selectins in mediating eosinophil migration to sites of inflammation in vivo has only been recently demonstrated in LPS mouse pleurisy model (Henriques et al. 1996). Our results in BCG pleurisy model have confirmed a role for L-selectin in eosinophil influx induced by bacterial agents. Taken together, our results have shown that mouse pleurisy induced by BCG is a useful model to study the inflammatory reaction that occurs during the primary infection with mycobacteria. This model should be helpful to improve our understanding of the role of leukocytes and cytokines in the delicate balance between strategies used by Mycobacteria to survive within a host and concomitant efforts of the host to kill it. This work was supported by grants from CNPq and CAPES (Brazil). References
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97193b.jpg] [oc97193a.jpg] [oc97193c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}