
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 92, Num. s2, 1997, pp. 233-235
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92 (Suppl.II), 1997, pp.
233-235
Nitric Oxide Mediates the Microbicidal Activity of Eosinophils
Sandra HP Oliveira, Simone G Fonseca, Pedro RT Romao, Sergio H Ferreira,
Fernando Q Cunha^+
Departamentos de Farmacologia e Microbiologia, Parasitologia e Imunologia,
Faculdade de Medicina de Ribeirao Preto, Universidade de Sao Paulo, Av.
Bandeirantes 900, 14049-900 Ribeirao Preto, SP, Brazil
^+Corresponding author. Fax: +55-16-633.2301. E-mail: fdqcunha@fmrp.usp.br
Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97194
Sizes of Files:
Text: 15.4K
Graphics: Line drawings and photographs (jpg) - 24.7K
There are several experimental evidences that nitric oxide (NO) is
involved in the microbicidal activity of macrophages against a number of
intracellular pathogens including Leishmania major, Trypanozoma cruzi,
Toxoplasma gondii. It is also well known that eosinophils (EO) have
microbicidal activity against many parasites such as Schistosoma
mansoni, Trichinella spiralis, T. cruzi and L. amazonensis. The
purpose of this study was to investigate if NO is involved in the
microbicidal activity of EO against L. major. Eosinophils harvested
from peritoneal cavity of rats released spontaneously after 24 and 48 hr a
small amount of nitrite. This release was enhanced by the treatment of
cells with IFN-gamma (200 IU/ml). This release was blocked by addition of the
NO synthase inhibitor, L-NIO (100 mM) into the culture. To determinate the
leishmanicidal activity of eosinophils the parasites were incubated with
activated eosinophils with IFN-gamma and the ability of surviving parasites to
incorporate [^3H]thymidine was evaluated. IFN-gamma-activated eosinophils were
able to kill L. major and to release high levels of nitrite. The
ability to destroy L. major and the release of NO were completely
blocked by L-NIO. These results indicate that activated eosinophils release
NO which is involved in the microbicidal activity of these cells against
L. major.
Key words: nitric oxide - eosinophils - microbicidal activity -
Leishmania major
Nitric Oxide Mediates the Microbicidal Activity of Eosinophils (Full
Text)
The microbicidal activity of eosinophils has been associated with
degranulation and release of granule-protein such as major basic protein
(MBP), eosinophil peroxidase (EPO), eosinophil cationic protein (ECP) and
eosinophil-derived neurotoxin (EDN). These substances are cytotoxic to
cells and to several parasites in vitro (Gleich et al. 1992), among
them, Trichinella spiralis (Wassom & Gleich 1979), Onchocerca
volvulus (Greene et al. 1981), Toxocara canis (Badley et al.
1987), Fasciola hepatica (Duffus et al. 1980), Necator
americanus (Desakorn et al. 1987), Nippo-strongylus brasiliensis
(Kojima et al. 1985), Schistosoma mansoni (Capron et al. 1979,
Butterworth et al. 1979), Trypanosoma cruzi (Villalta & Kierszenbaum
1984), Plasmodium falciparum (Waters et al. 1987), Leishmania
mexicana amazonensis (Pimenta et al. 1987) and L. donovani
(Pearson et al. 1987). In addition to the granule proteins, oxygen-free
radicals, including the toxic singlet oxygen, seem to contribute to the
microbicidal activity of eosinophils (Pincus et al. 1984).
Nitric oxide (NO) or nitrogen-derived metabolites have been identified as
major effector molecules involved in the macrophage microbicidal activity
against most intracellular pathogens, including L. major (Liew &
Millott 1990), Toxoplasma gondii (Adams et al. 1990), T.
musculi (Vincendeau & Dalouede 1991), T. cruzi (Gazzinelli et
al. 1992), P. berghei (Mellouk et al. 1991), Mycobacterium
leprae (Adams et al. 1991), M. avium (Denis 1991), Candida
albicans (Cenci et al. 1993) and the virus Ectromelia, vaccinia and
herpes simplex-1 (Gunasegaran et al. 1993). Recently, it was demonstrated
that the microbicidal activity of neutrophils against C. albicans is
also mediated by NO (Fierro et al. 1996).
Since at present there is no experimental demonstration of the contribution
of nitric oxide to the microbicidal activity of eosinophils, the aim of
this study was to investigate whether eosinophils produce nitric oxide and
whether NO is involved in the microbicidal activity of these cells against
L. major.
To investigate if eosinophils are able to produce nitric oxide, these cells
were harvested from peritoneal cavities of rats and incubated in vitro
with IFN-gamma. Eosinophils released spontaneously a small amount of
nitrite when incubated in vitro. Levels of nitrite in the medium
were already present after 12 hr of incubation, which increased
continuously within 48 hr. The stimulation of eosinophils with mIFN-gamma
enhanced the release of the nitrite after 24 and 48 hr of incubation. A
similar effect of IFN-gamma has been described in macrophages (Liew & Millott,
1990, Cunha et al. 1993). Nitrite production by the stimulated eosinophils
was abolished by the NO synthase inhibitor L-NIO^ (McCall et al. 1992),
indicating that the nitrite measured by the Griess method resulted from the
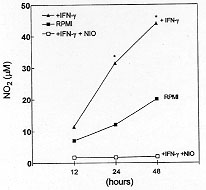
oxidation of L-arginine by NO synthase (Fig. 1).
Fig. 1: level of
nitrite in the culture supernatants of
eosinophils incubated with RPMI, mIFN-gamma (200 IU/ml) in the presence or
absence of L-NIO (100 mM). The nitrite concentration was determined at 12
hr , 24 hr and 48 hr after eosinophils stimulation by the Griess methods.
Data are reported as means +/- SEM of assays performed in triplicate and
are representative of two different experiments. * p < 0.05 compared to
RPMI (ANOVA followed by Bonferroni t-test).
An important protective role for reactive nitrogen intermediates has been
established in macrophage killing of intracellular protozoa (Liew & Millott
1990, Adams et al. 1990, Vincendeau & Dalouede 1991, Gazzinelli et al.
1992), bacteria (Adams et al. 1991, Denis 1991), fungus (Cenci et al. 1993)
and virus (Gunasegaran et al. 1993). A similar role has also been
demonstrated in neutrophil killing of C. albicans (Fierro et al.
1996) and Staphilococcus aureus (Malawista et al. 1992). Since there
are data showing that eosinophils are able to kill Leishmania
(Pimenta et al. 1987, Pearson et al. 1987), we investigated whether NO
mediates the killing of this intracellular parasite by stimulated
eosinophils. IFN-gamma-activated eosinophils were able to kill L. major
and to release high levels of nitrite. The ability to destroy L.
major and the release of NO were completely blocked by L-NIO,
suggesting that NO mediate the leishmanicidal activity of eosinophils.
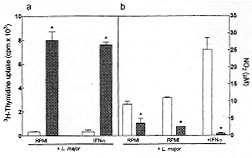
Leishmania survival inside the unstimulated eosinophils was not
affect by L-NIO treatment (Fig. 2). These results, together with the
findings that eosinophils are present in Leishmania lesions
(Katakura et al. 1993)^ , reinforce the importance to investigate the role
of eosinophils in the evolution of leishmaniasis.
Figure 2: killing of
Leishmania major by
eosinophils incubated with RPMI, or stimulated with mIFN-gamma (200 IU/ml) in
the presence or absence of L-NIO (100 mM) (panel a). Panel b: level of
nitrite in the supernatant of eosinophils incubated with L. major
and stimulated with IFN-gamma (200 IU/ml). Leishmanicidal activity is reported
as the ability of residual parasites to incorporate [^3H]thymidine. Data
are presented as means +/- SEM of assays performed in triplicate and are
representative of three different experiments. * p < 0.05 compared to RPMI
(ANOVA followed by Bonferroni t-test).
Thus, the data obtained in the present study indicate that activated
eosinophils release NO which may be involved in the microbicidal activity
of these cells against L. major.
REFERENCES
Adams B, Hibbs JB, Taintor RR, Krahenbuhl JL 1990. Microbiostatic
effect of murine activated macrophages for Toxoplasma gondii. Role
for synthesis of inorganic nitrogen oxides from L-arginine. J
Immunol 144: 2725-2729.
Adams LB, Franzblau SG, Vavrin V, Hibbs JRJ, Krahenbuhl JL 1991.
L-arginine-dependent macrophage effector functions inhibit metabolic
activity of Mycobacterium leprae. J Immunol 147: 1642-1646.
Badley JE, Grieve RB, Rockey JH, Glickman LT 1987. Immune mediated
adherence of eosinophils to Toxocara canis infective larvae: the
role of excretory-secretory antigens. Parasite Immunol 9:
133-143.
Butterworth AE, Wassom DL, Gleich GJ, Loegering DA, David JR 1979.
Damage to schistosomula of Schistosoma mansoni induced directly by
eosinophil major basic protein. J Immunol 122: 221-229.
Capron M, Torpier G, Capron A 1979. In vitro killing of S. mansoni
schistosomulo by eosinophils from infected rats: role of cytophilic
antibodies. J Immunol 123: 2220-2230.
Cenci E, Romani L, Mencacci A, Spaccapelo R, Schiabbella E, Paccetti P,
Bistoni F 1993. Interleukin-4 and interleukin-10 inhibit nitric
oxide-dependent macrophage killing of Candida albicans. Eur J
Immunol 23: 1034-1038.
Cunha FQ, Moss DK, Leal LMCC, Moncada S, Liew FY 1993. Induction of
macrophages parasiticidal activity by Staphylococcus aureus and
exotoxins through the nitric oxide synthasis pathways. Immunology
78: 563-567.
Denis M 1991. Tumor necrosis factor and granulocytes macrophages-colony
stimulating factor stimulate human macrophages to restrict growth of
virulent Mycobacterium avium and to kill avirulent M. avium:
Killing effector mechanism depends on the generation of reactive nitrogen
intermediates. J Leuk Biol 49: 380-387.
Desakorn V, Suntharasamai P, Pukrittayakamee S, Migasena S, Bunnag D
1987. Adherence of human eosinophils to infective filariform larvae of
Necator americanus in vitro, Southeast Asian. J Trop Med Pub
Health 18: 66-72.
Duffus WP, Thorne K, Oliver R 1980. Killing of juvenile Fasciola
hepatica by purified bovine eosinophil proteins. Clin Exp
Immunol 40: 336-340.
Fierro IM, Barja-Fidalgo C, Cunha FQ, Ferreira SH 1996. The involvement
of nitric oxide in the anti-Candida albicans activity of rat
neutrophils. Immunology 89: 295-300.
Gazzinelli RT, Oswald IP, Heiny S, James SL, Sher A 1992. The
microbicidal activity of interferon treated macrophages against
Trypanosoma cruzi involves an L-arginine-dependent, nitrogen
oxide-mediated mechanism inhibitable by interleukin-10 and transforming
growth factor-b. Eur J Immunol 22: 2501-2506.
Gleich GJ, Adolphson CR, Leiferman KM 1992. Eosinophils, p. 663. In JI
Galli, IM Goldestein, R Snyderman (eds). Inflammation: Basic Principles
and Clinical Correlates, 2^nd ed., Raven Press, New York.
Greene BM, Taylor HR, Aikawa M 1981. Cellular killing of microfilariae
of Onchocerca volvulus: eosinophil and neutrophil-mediated immune
serum-dependent destruction. J Immunol 127: 1611-1618.
Gunasegaran K, Xie Q-W, Buller RM, Nathan C, Duarte C, MacMicking JD
1993. Inhibition of viral replication by interferon-g-induced nitric oxide
synthase. Science 261: 1445-1447.
Katakura K, Saito S, Hamada A, Matsuda H, Watanabe N 1993. Cutaneous
leishmaniasis in mast cell-deficient W/W^v mice. Infect Immunity
61: 2242-2244.
Kojima S, Yamamoto N, Kanazawa T, Ovary Z 1985. Monoclonal
IgE-dependent eosinophil cytotoxicity to haptenated schistosomula of
Schistosoma japonicum: enhancement of the cytotoxixity and
expression of Fc receptors for IgE by Nippostrongylus brasiliensis
infection. J Immunol 34: 2719-2722.
Liew FY, Li Y, Millott S 1990. Tumor necrosis factor-a synergizes with
IFN-gamma in mediating killing of Leishmania major through the
induction of nitric oxide. J Immunol 145: 4306-4310.
Malawista SE, Montgomery RR, Van Blaricom G 1992. Evidence for reactive
nitrogen intermediates in killing of Staphylococci by human
neutrophil cytoplasts. A new microbicidal pathway for polymorphonuclear
leukocytes. J Clin Invest 90: 631-636.
McCall TB, Palmer RMJ, Moncada S 1992. Induction of nitric oxide
synthase in rat peritoneal neutrophils and its inhibition by dexamethasone
and the cytokine interleukin-8, p. 140. In S Moncada, MA Marletta, JB
Hibbs Jr, EA Higgs (eds). The Biology of Nitric Oxide: 2 Enzymology,
Biochemistry and Immunology, Vol 1, Portland Press, London.
Mellouk S, Green SJ, Nacy CA, Hoffman SL 1991. IFN-gamma inhibit
development of Plasmodium berghei exoerythrocytic stages in
hepathocytes by an L-arginine-dependent effector mechanism. J Immunol
146: 3971-3976
Pearson RD, Uydess IL, Chapman SW, Steigbigel RT 1987. Interaction of
human eosinophils with Leishmania donovani. Ann Trop Med
Parasitol 81: 735-739.
Pimenta PFP, Dos Santos MAV, De Souza W 1987. Fine structure and
cytochemistry of the interaction between Leishmania mexicana
amazonensis and rat neutrophils and eosinophils. J Submicrosc Cytol
19: 387-395.
Pincus SH, Butterworth AE, David JR, Robbins M, Vadas MA 1984.
Eosinophil-mediated killing of Schistosoma mansoni: oxidative
requirement for enhancement by eosinophil colony stimulating factor (CSFa)
and supernatants with eosinophil cytotoxicity enhancing activity (ECEA).
Cell Immunol 87: 424-433.
Villalta F, Kierszenbaum F 1984. Role of inflammatory cells in Chagas'
disease. I. Uptake and mechanism of destruction of intracellular
(amastigote) forms of Trypanosoma cruzi by human eosinophils. J
Immunol 132: 2053-2058.
Vincendeau P, Dalouede S 1991. Macrophages cytostatic effect on
Trypanosoma musculi involves an L-arginine-dependent mechanism.
J Immunol 146: 4338-4343.
Wassom DL, Gleich GJ 1979. Damage to Trichinella spiralis
newborn larvae by eosinophil major basic protein. Am J Trop Med Hyg
28: 860-863.
Waters LS, Taverne J, Tai PC, Spry CJ, Targett GA, Playfair JH 1987.
Killing of Plamodium falciparum by eosinophil secretory products.
Infect Immunity 55: 877-881.
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc97194b.jpg]
[oc97194a.jpg]
|

{kind=link}
{kind=link}