 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Does the Eosinophil Have a Protective Role in Amebiasis? Martha Lopez-Osuna/^+, Juan R Velazquez*, Roberto R Kretschmer
Immunology Research Unit, Scientific Research, CMN-SXXI, IMSS, P.O.Box
73-032, Mexico, D. F., 03020, Mexico ^+Corresponding author. Fax: This work was supported by a grant PCSABNA 0211552 from the National Council of Science and Technology of Mexico (CONACYT). Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97195
Sizes of Files:
Text: 14.4K
Graphics: Line drawings and photographs (jpg) - 35.7K
Tables (jpg) - 35.7K
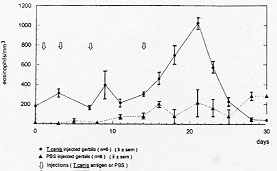
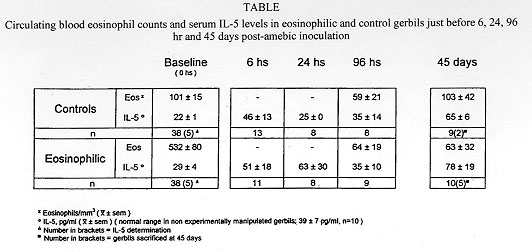
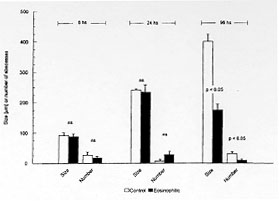
While normal human eosinophils are destroyed in vitro by virulent Entamoeba histolytica, notwhistanding the presence of antibodies and complement, activated eosinophils promptly destroy the parasite although dying also at the end of the process. To study the possible in vivo participation of eosinophils in invasive amebiasis, we compared the induction of experimental amebic abscess of the liver (AAL) in gerbils (Meriones unguiculatus) previously made eosinophilic through Toxocara canis antigen injection and in normal control gerbils. After intraportal inoculation of 10^5 ameba trophozoites (6 and 24 hr), the ratio of gerbils with AAL, as well as the number and size of the microabscesses was comparable in eosinophilic and control gerbils. However, at 96 hr the number and size of the microabscesses were significanly smaller (p<0.05) in eosinophilic gerbils. On the other hand the actuarial AAL survival curve up to 45 days post-amebic inoculation was signficantly (p<0.05) shifted to the right in controls. These results suggest that antigen-induced eosinophilia may exert a protective effect against AAL in gerbils. Key words: eosinophilia - Entamoeba histolytica - amebic abscess of the liver - gerbils Amebiasis is a major health problem in developing countries. Entamoeba histolytica infects 5 x 10^8 people worldwide, more than 90% of them living in West and South East Africa, China, SE Asia, Mexico, the western portion of South America and India (Smyth 1994). Only 10% of all infected individuals eventually develop invasive amebiasis ranging from mild (i.e. self-limited) or serious intestinal amebiasis, to life-threatening hepatic invasion (Walsh 1986). It has been suggested that there are in fact two different species of Entamoeba. One, the classic pathogenic E. histolytica discovered by Schaudinn in 1903. The other, the non-pathogenic E. dispar described by Brumpt in 1925, based mainly on epidemiological considerations. The concept of two Entamoeba species began to be objectively established by functional in vitro studies (Martinez-Palomo 1982, Mirelman & Chayen 1990) and zymodeme patterns (Sargeaunt 1988), culminating now with molecular genetic data (Tannich & Burchard 1991, Tannich 1996). Thus, the majority of ameba infected individuals may actually harbour E. dispar, leaving the E. histolytica as the agent of the relatively rare invasive intestinal and extraintestinal disease. Amebic abscess of the liver (AAL) on the other hand, occurs in less than 1% of individuals with invasive intestinal amebiasis (Martinez-Palomo et al. 1993). Why the vast majority of individuals (=/>99%) appear to be resistant to extraintestinal invasion remains unknown but some inroads have now been made: a significant increase in the SCO1 complotype and in HLA-DR3 was found in 160 Mexican mestizos (adults and children) with AAL when compared to healthy controls, to adult patients with amebic rectocolitis or to asymptomatic amebic carriers (Arellano et al. 1996). DNA subtyping is now in process to confirm these findings. Studies of human amebiasis and of experimental animal models of amebic disease have suggested that cell-mediated rather than humoral immune mechanisms, are responsible for acquired immunity to AAL (De Leon 1970, Salata & Radvin 1986, Kretschmer & Lopez-Osuna 1990). Moreover, axenically grown trophozoites of E. histolytica kill in vitro normal human neutrophils and eosinophils (not-withstanding the aid by antibody and complement), normal monocytes and lymphocytes, without suffering changes in their own viability (Jarumilinta & Kradolfer 1964, Artigas et al. 1966, Guerrant et al. 1981, Salata et al. 1985, 1987, Lopez-Osuna et al. 1986, Lopez-Osuna & Kretschmer 1989). However, if macrophages are activated beforehand, they become capable of effectively destroying virulent amebas, even though finally succumbing in the process as well (Salata et al. 1985). In our laboratory we found that the same occurs with activated human peripheral blood eosinophils (Lopez-Osuna et al. 1992). There is no circulating eosinophilia in most protozoan infections, yet this is not necessarily an argument against their possible participation in AAL, specially because eosinophils are regularly found in the early inflammatory stages of experimental amebic liver disease (Tsutsumi et al. 1984), and since the bulk of eosinophils in the body normally dwells, not intravascularly but in tissues (Weller 1991). We therefore decided to study in vivo the role of eosinophils, by inducing eosinophilia in gerbils (M. unguiculatus) through four injections of an innocuous Toxocara canis antigen, prior to the induction of experimental AAL with 1 x 10^5 intraportally injected ameba trophozoites (Velazquez et al. 1995, Lopez-Osuna 1995, Velazquez 1996). This relatively small inoculum of amebas (compared to the usual 1 x 10^6) was chosen in order to maximize the eosinophil's chances to reveal its eventual protective role against E. histolytica. Gerbils reached their eosinophilia peak at 21 days (after the first injection of the antigen) (Fig.1 and Table) at which time they and their controls (i.e.normal eosinophil number) received the intraportal injection of E. histolytica HM1-IMSS in saline. Animals were then^ sacrificed at 6, 24, 96 hr and - those surviving - at 45 days postinoculation.
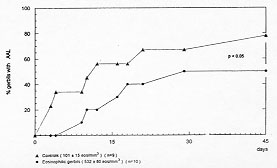
Figure 3: actuarial curve of eosinophilic gerbils with AAL. Cumulative percent of eosinophilic (solid circle) and control (solid triangle) gerbils dying spontaneously with ambic abscess of the liver within 45 days.
In order to confirm these findings, we are now studying AAL in eosinopenic gerbils. A search is also presently under way for specific eosinophil granule proteins (MBP, ECP, EPO) in the livers of eosinophilic gerbils in order to ascertain the true presence or absence of this leucocyte (Gleich et al. 1994). Much remains to be learned in order to establish if eosinophils actually play a role in invasive amebiasis. Screening carefully those gerbils that escape (or recover) from the formation of amebic abscesses (rather than studying only those with AAL) appears now to be mandatory. The traditionally eschewed subject of active participation of eosinophils in protozoan diseases (trypanosomiasis, malaria and leishmaniasis) has become an important landmark in today's immunoparasit-ology. Invasive amebiasis could apparently join this group. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97195c.jpg] [oc97195d.jpg] [oc97195a.jpg] [oc97195b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}