 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Serotypes of Vibrio cholerae Non-O1 Isolated from Water Supplies for Human Consumption in Campeche, Mexico and their Antibiotic Susceptibility Pattern
Angelica P Isaac-Marquez/^+, Claudio M Lezama-Davila*, Carlos Eslava-
Centro de Investigaciones en Enfermedades Tropicales, Universidad Autonoma
de Campeche, Av. Agustin Melgar s/n, 24030, Campeche, Mexico Received 5 March 1997; Accepted 27 August 1997
Code Number:OC98004
Sizes of Files:
Text: 25.7K
Graphics: Tables (jpg) - 68.3K
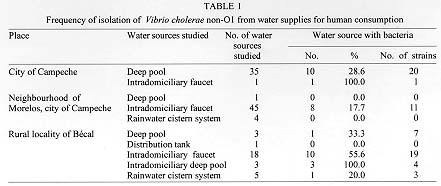
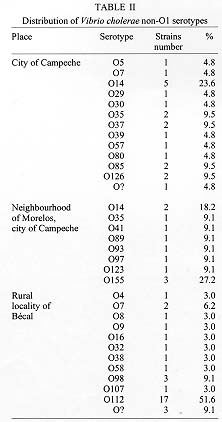
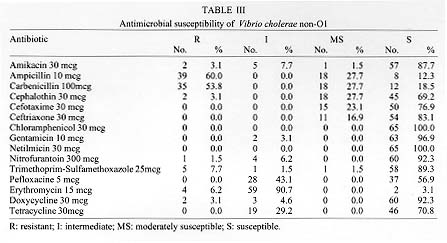
The presence of Vibrio cholerae non-O1 in water supplies for human consumption in the city of Campeche and rural locality of Becal was investigated. V. cholerae non-O1 was detected in 5.9% of the samples obtained in deep pools of Campeche. Studies conducted in Becal and neighbourhood of Morelos in Campeche indicated that collected samples harbored V. cholerae non-O1 in 31.5% and 8.7% respectively. There was a particular pattern of distribution of V. cholerae non-O1 serotypes among different studied regions. Accordingly, V. cholerae non-O1 serotype O14 predominated in the deep pools of Campeche and together with V. cholerae non-O1, O155 were preferentially founds in samples taken from intradomiciliary faucets in the neighbourhood of Morelos. Samples from Becal predominantly presented the serotype O112. 60% and 53.8% of all studied strains of V. cholerae non-O1 proved to be resistant to ampicillin and carbenicillin. 3.1%, 7.7% and 6.2% presented resistant to doxycycline, trimethoprim-sulfamethoxazole and erythromycin respectively. The study showed the necessity of performing a strong epidemiologic surveillance for emergence and distribution of V. cholerae non-O1. Key words: Vibrio cholerae non-O1 - serotypes - antibiotic susceptibility - water supplies - Campeche - Mexico Vibrio cholerae serogroup non-O1 are autochthonous bacteria of aquatic environments (Morris 1990) and right from the 1950s and 1960s has been identified in outbreaks of gastroenteritis in different parts of the world (McIntyre et al. 1965, Aldova et al. 1968, El-Shawi & Thewaini 1969, Kamal 1971). Currently it is recognized that non-O1 serogroup includes approximately 150 serotypes and play an important role as an aethiological agent of gastrointestinal illness ranging from mild watery diarrhea to febrile enteritis with bloody diarrhea. In contrast to V. cholerae O1, V. cholerae non-O1 causes extraintestinal infections (Sanyal 1992); but gastroenteritis is the most common clinical manifestation (Morris 1990, WHO 1993). In October 1992, an epidemic of cholera-like illness caused by a V. cholerae serogroup non-O1 broke in Madras, India. The causative agent was found to be a new serogroup of V. cholerae, defined as O139, with the synonym Bengal, in recognition of the origin of this strain (Shimada et al. 1993, Ramamurthy et al. 1993). This strain has also been responsible for epidemic outbreaks of cholera in Bangladesh, southern Asia and U.S.A. (Albert et al. 1993, Bhattacharya et al. 1993, CDC 1993, Gonzalez et al. 1993) with possible disemination to Latin America throughout imported cases.^ Before of this epidemics of V. cholerae non-O1, O139 only V. cholerae O1 was considered capable of producing epidemics of cholera. In Peru, cholera appeared in January 1991 and spreaded quickly in Latin America (PAHO 1995). The disease evolved in explosive epidemics, the largest recorded since the beginning of the seventh pandemic in Indonesia in 1961. The first reported cases of cholera in the state of Campeche, located in southeast of Mexico, were registered in August 1991 and because of a number of environmental conditions determined by economic, sociologic and educational factors, this disease spread quickly and from 1992 became one of the federal states with more reported cases of this disease, registering the first places in morbility and mortality rates in Mexico (DGE 1993, 1995, Valdespino 1994). From 1992 up to now city of Campeche has experienced several outbreaks of cholera and from July to September, 1993 Becal, rural locality of Campeche, registered an important number of cases of this disease. The purpose of this study was the characterization of serotypes and the antibiotic susceptibility pattern of V. cholerae non-O1 strains isolated from water supplies for human consumption before, during and after those outbreaks of cholera. MATERIALS AND METHODS This study was performed in three different steps. The first one was from August 1992 to July 1993 and included seven samplings of water coming from 35 deep pools that provide water to the city of Campeche. The second step was carried out during August to December 1993 and included two collections of samples from the distribution network placed in the rural locality of Becal. In addition, were examined intradomiciliary deep pools and rainwater cistern systems which are commonly used in this region for human consumption. The third step was performed during July 1994 to February 1995 and included two water monitorings of neighbourhood of Morelos in the city of Campeche. Testing of total residual chlorine concentration - Presence of total residual chlorine in water samples was evaluated by the ortotoluidine method (Rodier 1990). Chlorine concentration studied varied from 0 to 3 ppm. Isolation - The Spira swab (Spira & Ahmed 1981) and double enrichment in alkaline peptone water (Chandra 1992) methods were used for the isolation of V. cholerae. In both cases were employed the TCBS agar (thiosulfate-citrate-bile-sucrose agar). The water samples were storaged and transported at 4 C and processed within the next 2 hr of their collection. Identification - Yellow colonies from TCBS agar were submitted to the standard series of biochemical media used for identification of members of the Vibrionaceae family (Holt et al. 1994). The serological identification was determined by slide agglutination test using polyvalent O1 and type-specific Inaba and Ogawa antisera (Difco) and by coagglutination test employing monoclonal antibody anti-V.cholerae O1 adsorbed to heat-killed S. aureus (New Horizons Diagnostics Co.). V. cholerae serogroup O1 and non-O1 were used as controls. Serological classification of V. cholerae non-O1 - Polyvalent antisera for serological classification of different V. cholerae non-O1 serotypes were used. The reference strains of V. cholerae non-O1 serotypes O1 to O83 were kindly provided by Dr Bernard Rowe, Head of Enteric Pathogens Central Laboratory of Public Health, London, England, and those reference strains from serotype O84 to O140 and O155 were provided by Dr T Shimada from the National Institute of Health, Tokyo, Japan. Antigens used for serological classification were obtained by heating bacterial cultures at 100^ C for 1 hr and adjusted with tube 3 of McFarland nephelometer; 1:100 serum dilutions were used in a proportion of antigen/sera (V/V) of 50 ml in 96 wells microtitration plates, incubated at 50 C for 24 hr. Sera that agglutinated with this antigen preparation was serially diluted twice to know the titer and so determinate identity of each serotype (Sakazaki & Donovan 1984, Shimada et al. 1994).^ Antimicrobial susceptibility testing - All V. cholerae non-O1 isolates were tested for antibiotic susceptibility to 15 antibiotics (Difco) by Kirby-Bauer method (NCCLS 1988) and strains were recorded as susceptible, intermediate, moderately susceptible or resistant. RESULTS The results indicate that in 41.1% (14/34) and 80.7% (67/83) of the samples collected from intradomiciliary faucets of rural locality of Becal and neighbourhood of Morelos a range of 0.5-1.5 ppm and 0.3-3 ppm of total residual chlorine was detected, respectively, by the ortotoluidine method. In the case of deep pools in Morelos and Becal, 76.9% (10/13) and 12.5% (1/8) of samples collected presented a range of 1-3 ppm. Samples obtained both from deep pools in Campeche and from the distribution tanks, intradomiciliary deep pools and rainwater cistern systems in the studied areas did not contained chlorine. In Table I it is shown the frequency of isolation of V. cholerae non-O1 strains in different water sources. 5.9% (13/222) of water samples taken from deep pools of the city of Campeche, and 31.5% (17/54) and 8.7% (9/104) of those water samples from Becal and Morelos respectively harbored V. cholerae non-O1. Biochemical features of isolated strains were very similar to those reported by Holt et al. (1994). It has been reported that V. cholerae do not perform esculin hydrolysis, however, 67.7% (44/65) of those strains isolated can indeed to hydrolize this compound at 0 and 1% of sodium chloride. In addition, 3.1% (2/65) of bacterial strains grew in nutrient agar with 8% of NaCl. A total of 65 strains belonging to 29 different serotypes were identified during the period of study. In Table II it is shown the distribution and frequency of V. cholerae non-O1 serotypes in relation to the total of bacterial strains isolated in the studied regions. The 35.7% (5/14) of samples analyzed from deep pools in Campeche contained V. cholerae non-O1 serotype O14. In the neighbourhood of Morelos 33.3% (3/9) and 22.2% (2/9) of samples taken from intradomiciliary faucets harbored V. cholerae serotypes O155 and O14 respectively. Only in one out of three deep pools of Becal was isolated V. cholerae non-O1 serotype O112 in all the tested samples. Additionaly, in 66.7% (2/3) of those samples were isolated bacterias that biochemically behave as V. cholerae non-O1 but did not agglutinate with any of the 141 anti-V. cholerae non-O1 polyvalent sera (anti-V. cholerae non-O1 serotype O1 to O140 and O155) availables. In 80% (8/10) of tested samples of intradomiciliary faucets was detected V. cholerae non-O1 serotype O112. The water samples taken from the intradomi-ciliary deep pools in Becal registered the presence of V. cholerae non-O1 serotypes O58, O38, O8 and O4. It was demonstrated the presence of V. cholerae non-O1 serotype O98 in rainwater cistern systems. The pattern of susceptibility of 65 isolated strains of V. cholerae non-O1 with 15 different antibiotics is shown in Table III. More than 67.7% (44/65) of the isolated strains were susceptible to the aminoglycosides, cephalosporins, tetracyclines, chloramphenicol, nitrofurantoin and trimethoprim-sulfamethoxazole. In contrast, 50.8% (33/65) of all strains showed resistance to two or more drugs. 49.2% (32/65) of these microorganisms were resistant to ampicillin and carbenicillin, while 10.8% (7/65) were resistant only to ampicillin and 4.6% (3/65) exhibited resistance only to carbenicillin. The rate of resistance to these substances was higher than to the other drugs. In addition, 3.1% (2/65), 7.7% (5/65) and 6.2% (4/65) of V. cholerae non-O1 strains presented resistance to the doxycycline, trimethoprim-sulfamethoxazole and erythromycin respectively. DISCUSSION Based in the results of the present study it is possible to conclude that the water distribution system plays an important role as transmission vehicle for V. cholerae non-O1. Considering the presence of V. cholerae non-O1 in the studied sources, it is possible to assume that this organism might be responsible of a proportion of diarrheal diseases among the population of the city of Campeche and the rural locality of Becal. It is important to observe that 29 V. cholerae non-O1 serotypes found in this study display a clear pattern of geographic distribution. As can be observed in the Table II V. cholerae non-O1, O112 was located only in Becal, and V. cholerae non-O1, O14 and O155 were exclusively located in the city of Campeche. Regarding to those serotypes with a less frequent presentation; those present in Becal are different to the ones found in the city of Campeche with only one exception that corresponded to V. cholerae non-O1, O7. V. cholerae non-O1, O14 and O35 were present in samples taken from city of Campeche and neighbourhood of Morelos, nonetheless the serotypes of the remaining strains were different. Additionaly, 9.1% (3/33) and 4.8% (1/21) of those strains isolated in Becal and the city of Campeche, respectively belong to a particular V. cholerae non-O1 strain which do not corresponded to any of those 141 serotypes that were studied in this work and might represent an autoctonous uncharacterized serotypes. The presence of 2 or 3 different serotypes of V. cholerae non-O1 were detected in 21.4% (3/14), 22.0% (2/9) and 29.4% (5/17) of samples collected from the city of Campeche, neighbourhood of Morelos and the rural locality of Becal respectively. This indicate that coexistance of different serotypes of V. cholerae non-O1 frequently happens within the same habitat. Nonetheless that a high percentage of strains isolated during the development of this study displayed susceptibility to a diverse group of antibiotics, it is important to observe that V. cholerae non-O1 strains presented an important resistance to doxycycline, trimethoprim-sulfamethoxazole and erithromycin. Additionaly, a hight percentage of those strains registered an intermediate susceptibility to erithromycin and tetracycline. This susceptibility pattern represent a significant information because they are the antibiotics that WHO recommends for the treatment of cholera. In Mexico during 1991, 3.2% of clinical isolates of V. cholerae O1 presented resistance to tetracycline and 1.5% to doxycycline. During 1992 this multiresistance decreased to 1.4% and from 1993 to 1995 only 0.1% of V. cholerae O1 strains registered resistance to the antibiotics mentioned; nonetheless the strains isolated conserved their resistance to ampicillin (Gutierrez-Cogio 1995). In Mexico, there is not available information about antibiotic susceptibility pattern of environmental isolates of V. cholerae non-O1. However, according to the results obtained in this study it is possible to conclude that the environmental V. cholerae non-O1 strains are significantly more resistant to antibiotics than the clinical V. cholerae O1 strains. This acquired an important significance, because of phenomenon of acquired plasmid-mediated resistance that might occur in a habitat where V. cholerae serogroup O1 and non-O1 coexists. Studies performed in Tanzania and Bangladesh reported strains of V. cholerae with plasmid-mediated resistance to a wide range of antibiotics (Mhalu et al. 1979, Threlfall & Rowe 1982). Dalsgaard et al. (1995) reported that of 23 environmental V. cholerae non-O1 strains, 78.3% exhibited resistance to tetracycline and 8.7% were resistant to ampicillin and chloramphenicol. These results differ from the results of the present study. Nevertheless, the resistance to trimethoprim/sulfamethoxazole and erythromycin were similar in both studies. It is possible that variations in the antibiotic susceptibility pattern could be connected with the origin and habitat conditions of the V. cholerae non-O1 strains and with a variety of antibiotics used in differents parts of the world. The sudden emergence of V. cholerae non-O1, O139 as ethiologic agent of epidemic cholera represents an important shift in the epidemiology of this disease. The evidence suggests the V. cholerae O1 El Tor gave rise to O139 by adquisition of novel DNA (Colwell 1996). Studies have shown that DNA sequence that determine the antigenic properties of the O139 cell surface is also present in two V. cholerae non-O1 strains with serotypes O69 and O141 (Colwell 1996) and additionaly that V. cholerae non-O1 serotypes O22 and O155 posses somatic (O) antigen factors in common with O139 strain (Mukhopadhyay et al. 1995). Cravioto et al. (1994) proved that the Bengal strain is genetically related to V. cholerae O1 El Tor Ogawa isolated in Mexico, and two other non-O1 strains (serotypes O22 and O30). In the present study V. cholerae serotypes O30 and O155 were detected in samples analyzed in the city of Campeche. Considering these finding, it is not possible to establish a line of division between V. cholerae O1, O139 and the bacterial strains belonging to serogroup non-O1. It is now evident that these microorganisms have acquired an increasing importance, particularly in relation to medical relevance and ecological distribution. The fact that the serogroup O139 emerged as responsible of epidemic cholera raised the possibility that such an event can occur once again, in relation to V. cholerae non-O1 serotypes prevalents in the Americas. It is recommended a strong epidemiologic surveillance in particular regarding emergence and distribution of V. cholerae non-O1 strains both in environmental samples and clinical specimens. Additional support to these study is found in the fact that V. cholerae non-O1 has been isolated in patients with diarrhoea in Mexico (Gutierrez-Cogio 1995). In addition, the relatively high prevalence of multiple antibiotic resistance of V. cholerae non-O1 strains recovered from water samples indicate the necessity of performing a epidemiologic surveillance of the pattern of antibiotic susceptibility. ACKNOWLEDGEMENTS To health authorities and the government of Campeche for the assistance provided during the development of this study. To the staff of the University of Campeche for technical assistance. This work was finantially supported by Secretaria de Educacion Publica (C 91-01-04-002-441, P/FOMES 93-04-01 and P/FOMES 94-04-03-01), the National University of Mexico and the University of Campeche. REFERENCES Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98004b.jpg] [oc98004a.jpg] [oc98004c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}