 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Estimation of Genetic Divergence and Gene Flow between Culex pipiens and Culex quinquefasciatus (Diptera: Culicidae) in Argentina Silvia G Humeres, Walter R Almiron*, Marta S Sabattini, Cristina N Gardenal/^+
Catedra de Quimica Biologica, Facultad de Ciencias Medicas, cc.35, suc.16,
5016 Cordoba, Argentina
Received 20 December 1996; Accepted 16 October 1997
Code Number:OC98011
Sizes of Files:
Text: 28.3K
Graphics: Line drawings and photographs (jpg) - 25.5K
Tables (jpg) - 112.9K
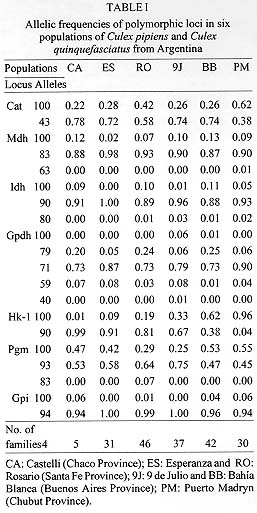
Allele frequencies at seven polymorphic loci controlling the synthesis of enzymes were analyzed in six populations of Culex pipiens L. and Cx. quinquefasciatus Say. Sampling sites were situated along a north-south line of about 2,000 km in Argentina. The predominant alleles at Mdh, Idh, Gpdh and Gpi loci presented similar frequencies in all the samples. Frequencies at the Pgm locus were similar for populations pairs sharing the same geographic area. The loci Cat and Hk-1 presented significant geographic variation. The latter showed a marked latitudinal cline, with a frequency for allele b ranging from 0.99 in the northernmost point to 0.04 in the southernmost one, a pattern that may be explained by natural selection (FST = 0.46; p < 0.0001) on heat sensitive alleles. The average value of FST (0.088) and Nm (61.12) indicated a high gene flow between adjacent populations. A high correlation was found between genetic and geographic distance (r = 0.83; p < 0.001). The highest genetic identity (IN = 0.988) corresponded to the geographically closest samples from the central area. In one of these localities Cx. quinquefasciatus was predominant and hybrid individuals were detected, while in the other, almost all the specimens were identified as Cx. pipiens. To verify the fertility between Cx. pipiens and Cx. quinquefasciatus from the northern- and southernmost populations, experimental crosses were performed. Viable egg rafts were obtained from both reciprocal crosses. Hatching ranged from 76.5 to 100%. The hybrid progenies were fertile through two subsequent generations. Key words: Culex pipiens - Culex quinquefasciatus - genetic divergence - gene flow - crossing experiments - Argentina Mosquitoes of the Culex pipiens L. Group are important vectors of St. Louis encephalitis (SLE) virus in the east-central United States and eastern Texas (Mitchell et al. 1980). A strain of SLE virus was isolated from Cx. pipiens quinquefasciatus Say collected in Santa Fe Province, Argentina (Mitchell et al. 1985), and a colony established from the same site, showed to be an efficient vector of both the Argentine and United States SLE viral strain (Mitchell et al. 1985). The members of this group are widely distributed throughout the world, with two predominant species: Cx. pipiens and Cx. quinquefasciatus. The former is restricted to cooler areas while the later inhabits warmer tropical and subtropical regions (Mattingly 1951, Laven 1967, Barr 1982). Morphological, ethological and ecological features have been used to characterize these species; however, their taxonomic status is still undefined in some geographic areas. Male genitalia morphology and morphometry (DV/D ratio) have been the main traits used to distinguish Cx. pipiens from Cx. quinquefasciatus. However, intergradation zones have been definited in North America (Sundararaman 1949, Barr 1957, Cheng 1976, Jakob et al. 1980, Barr 1982, Tabachnich & Powell 1983), northern Japan (Edwards 1921, Bekku 1956, Sasa et al. 1963, Ishii 1980), southeastern Australia (Dobrotworsky 1967, Barr 1982), and the Middle East (Edwards 1921, Harbach 1985, 1988). In Africa, the two taxa occur in sympatry but do not hybridize (Jupp 1978, Donaldson 1979). In Argentina (South America), hybrid forms have been found in a central area of the country between 30 degrees and 33 degrees S (Brewer et al. 1987, Almiron et al. 1995). Several authors have stressed the contribution of multilocus electrophoretic analysis to the knowledge of the genetic structure of populations of pipiens and quinquefasciatus from different countries and to clarify their taxonomic position (Cheng 1976, Tabachnick & Powell 1983, Urbanelli et al. 1985, Villani et al. 1986). Studies of genetic polymorphism have not been undertaken in the Cx. pipiens Group of South America. This paper presents a study of the distribution of allele frequencies for seven polymorphic enzyme loci in six populations of Cx. pipiens and Cx. quinquefasciatus from Argentina, situated along a north-south line of about 2,000 km, and an estimation of the degree of gene flow and genetic divergence among them. Crossing experiments between mosquitoes from the two extreme populations of this line were also performed. MATERIALS AND METHODS
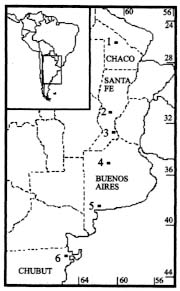
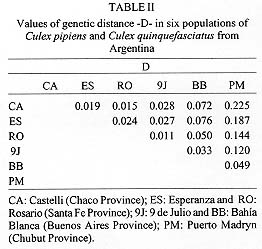
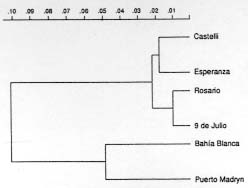
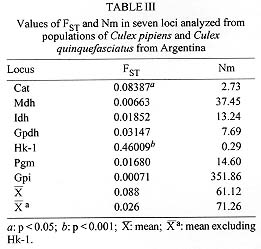
Figure 1 : location of Culex pipiens and Cx. quinquefasciatus collection sites: 1. Castelli; 2. Esperanza; 3. Rosario; 4. 9 de Julio; 5. Bahia Blanca; 6. Puerto Madryn.Females were collected from natural resting sites using baited (chicken) can traps, and then transported alived to the laboratory. In order to obtain egg rafts, the field females were placed individually in plastic tubes with wet cotton and filter paper at the bottom. The egg rafts were reared as individual progenies in plastic trays (25 +/-3 C, 14:10 L/D photoperiod). Larvae were fed with commercial rodent food until adults emerged. Adults were frozen at -40 C, 48 hr after emergence, until electrophoresis was performed. Five to seven males from each progeny were examined for phallosome morphology and morphometry (DV/D ratio) (Sundararaman 1949). Electrophoretic analysis - Homogenates were obtained from single individuals crushed in distilled water. The resulting suspension was absorbed in a Whatman 3 MM paper wick (2.5 mm) and inserted in starch gel trays. Homogenates from six to nine descendants of each progeny were placed in a gel block which admitted up to sixteen insertions. The number of families analyzed from each locality are indicated in Table I. Standard horizontal electrophoresis was carried out at 4 C. Gels were sliced longitudinally into four slabs and each one stained for specific enzymes (Humeres et al. 1990). The following buffer systems were used: continuous Tris-boric EDTA, pH: 8.6 (Markert & Faulhaber 1965), for the separation of catalases (CAT), glycerophosphate dehydrogenase (GPDH) and hexokinase (HK); discontinous Tris citrate, pH: 6.7 for gels and pH: 6.3 for the electrode cells (Shows & Ruddle 1968) for malate (MDH) and isocitrate (IDH) dehydrogenases, and continuous lithium hydroxide pH:8.4 (Steiner & Joslyn 1979) for phosphoglucomutase (PGM) and glucosephos-phate isomerase (GPI). Alleles were assigned a number according to the relative mobility of the proteins, using 100 as the protein migrating fastest toward the anode. The index of genetic distance (D) proposed by Nei (1972) was used to estimate the degree of genetic divergence between populations. The analysis was performed using the Genind program (Vilardi 1992). Gene flow was estimated using Wright's (1965) F-statistics as modified by Nei and Chesser (1983). Effective migration rate (Nm) was calculated from F[ST]. Crossing experiments - The following crosses were performed as indicated in Almiron et al. (1995): females Cx. pipiens (Puerto Madryn) x males Cx. quinquefasciatus (Castelli) and its reciprocal. RESULTS According to the analysis of male genitalia (DV/D ratio) the following species were identified: Cx. quinquefasciatus was exclusively present in the northern localities of Castelli and Esperanza. In Rosario, although Cx. quinquefasciatus was predominant, hybrids were also detected. Almost all individuals from the 9 de Julio sample belonged to Cx. pipiens except one Cx. quinquefasciatus. In the southern localities (Bahia Blanca and Puerto Madryn), only Cx. pipiens (Almiron et al. 1995) has been identified. Electrophoretic analysis - Zymograms obtained allowed the analysis of seven loci: Cat, Mdh, Idh, Gpdh, Hk-1, Pgm, and Gpi. Criteria of genetic interpretation was similar to those applied by other authors to polymorphisms detected in different mosquito species (Steiner & Joslyn 1979, Villani et al. 1986). Parental genotypes for each isofemale line were determined when unequivocal, and allelic frequencies were calculated. Data are summarized in Table I. In all cases, genotypes were distributed according to the Hardy-Weinberg equilibrium. The predominant alleles at the Mdh, Idh, Gpdh and Gpi loci presented similar frequencies in all samples. For the Pgm locus, allele 93 was predominant in Castelli and Esperanza, while allele 100 was the most frequent in Bahia Blanca and Puerto Madryn. The intermediate localities of Rosario and 9 de Julio presented similar frequencies. Allelic frequencies of Cat and Hk-1 differ markedly between extreme populations. Hk-1 shows a striking latitudinal cline. Values of Nei's genetic distance between the populations are presented in Table II. Samples including typical Cx. pipiens (Bahia Blanca, 9 de Julio and Puerto Madryn) show a low genetic distance. Samples from the localities of Castelli, Esperanza and Rosario, where Cx. quin-quefasciatus was predominant, were also genetically homogeneous. However, 9 de Julio and Rosario samples showed the lowest value of genetic distance (DN = 0.011), although they were represented almost exclusively by Cx. pipiens and Cx. quinquefasciatus respectively. Populations from extreme sites of the study area presented the highest value of genetic distance (D = 0.225). Fig. 2 summarizes these data in a dendrogram based on D values and clustered using the Genind program. A significant correlation between genetic and geographical distances was detected (r = 0.83; p < 0.001). In Table III, FST and Nm values are presented for the analyzed loci in the six populations. Figure 2: phenogram constructed using the unweighted pair group methods with arithmetic mean, based on Nei's genetic distance values among samples of Culex pipiens and Culex quinquefasciatus. Crossing experiments - Viable eggs were obtained from both crosses. Crosses involving female Cx. pipiens from Puerto Madryn x male Cx. quinquefasciatus from Castelli showed hatching rates of 100%, and 76.5% in the reciprocal. The F1 and F2 offspring of both crosses proved to be fertile, with hatching rates between 95 and 100% (Table IV). The colony was discarded after obtaining the F3.
DISCUSSION Geographic patterns of allele frequencies in Cx. pipiens and Cx. quinquefasciatus were analyzed for populations from different collection sites in an extended region of Argentina, including Esperanza where Cx. quinquefasciatus was found naturally infected with the St. Louis encephalitis virus (Mitchell et al. 1985). Although allele frequencies at one of the seven loci analyzed (Hk-1) showed a marked clinal gradient across the studied area (Table I), the lack of fixed alternative allozymes at any of the loci did not allow the characterization of Cx. pipiens and Cx. quinquefasciatus. Similar results were obtained by Cheng et al. (1982) and Tabachnick and Powell (1983). Hexokinase (Hk-1), 6-phosphogluconate dehydrogenase (6Pgdh), glucosephosphate isomerase (Gpi) and phosphoglucomutase (Pgm), exhibit a latitudinal cline in gene frequencies across the range of the species in the United States (Cheng et al. 1982). Tabachnick and Powell (1983) reported that the same loci track a topographical temperature gradient in the Central Valley of California. In our study, a similar pattern was found only for locus Hk-1, with a frequency for allele 100 ranging from 0.01 in the northernmost point of the sampling area, to 0.96 in the southernmost collection site (Table I). Alleles at this locus may be equivalent to those designated HkA and HkB by Cheng et al. (1982) for populations from North America, where HkA decreases in frequency with increasing latitudes. Allele frequencies at Pgm locus did not show correlation with DV/D ratios, as demonstrated by Cheng et al. (1982) and Tabachnick and Powell (1983). These alleles presented a particular distribution (Table I), being similar for populations from the warm and cold areas (Pgm 93: 0.53 and 0.58 for Castelli and Esperanza; 0.47 and 0.45 for Bahia Blanca and Puerto Madryn) and between the two intermediate ones (Pgm 93: 0.64 and 0.75 for Rosario and 9 de Julio). Population pairs with similar allele frequencies, share similar environmental conditions (not only temperature), since they occupy approximately the same geographic area. This may produce a non random distribution of genetic variants. An unique allele (c) for the Pgm locus was found in the sample from Rosario with a frequency of 7%. In this site, hybrid specimens (according to the DV/D ratio) were detected in a proportion of 4.7% (Almiron et al. 1995). The presence of rare alleles in hybrid populations has been reported for different animal groups; intragenic recombination was proposed to explain it (Golding & Strobeck 1983). However, we did not determine if the unique allele was confined to the hybrid specimens. Estimation of the standardized variance in genic frequencies among populations (FST) gave an average value of 0.088, and an average Nm of 61.12 (Table II) which would indicate that a high gene flow is occurring between adjacent populations according to Wright (1978). However, if Hk-1 is excluded, these values are 0.026 and 71.26 respectively. As it has already been pointed out, different alleles at the Hk-1 locus are present in high frequencies in populations from the extreme points (Hk-1 90: 0.99 in Castelli and 0.04 in Puerto Madryn; FST = 0.46, p < 0.0001) suggesting that natural selection at this locus is strong enough in preventing gene flow to homogenyze gene frequencies. Pryor (1980) demonstrated differential heat stability of different allele products at Hk-1 locus, being the heat-stable ones more common in Cx. quinquefasciatus. This finding would indicate that temperature could be an important factor determining genetic composition of populations at Hk-1 locus. Although in a lower degree, the locus Cat also showed significant local differentiation (FST = 0.08, p < 0.05), but there was no correlation with the sample origin. Another approach to understand patterns of geographic variation is the analysis of genetic distance values among populations. The lowest distance (Table III) correspond to the geographically closest samples (9 de Julio and Rosario, DN = 0.011) suggesting that Cx. pipiens and Cx. quin-quefasciatus are a stable interbreeding unit in those areas where temperature allows their survival. On the other hand, a good correlation between genetic and geographical distance (r = 0.83; p < 0.001) was found. It is difficult to explain this observation as produced by a drift-migration interaction, taking into account the FST values for different loci. It could be assumed that this correlation would be mainly determined by loci under the control of natural selection, being temperature an important selective factor for the maintenance of genetic cohesion among populations of the same species. The average value of D was 0.102, lower than that reported by Urbanelli et al. (1985) between Cx. pipiens from Italy and Cx. quinquefasciatus from Africa (D = 0.145). D values ranging from 0.05 to 0.2 are considered to correspond to populations of the same species (Ayala 1975). Almiron et al. (1995) performed different crosses between Cx. pipiens and Cx. quinquefasciatus from a central area of Argentina including three localities (Bahia Blanca, Buenos Aires Province; Cordoba, Cordoba Province; and Rosario, Santa Fe Province). Hatching from 70 to 100% was reported by those authors, except for one cross involving female Cx. pipiens from Bahia Blanca x male Cx. quinquefasciatus from Rosario, where a high incompatibility was observed (11.1% hatch). According to this situation, a low hatching rate was expected for the cross between female Cx. pipiens from Puerto Madryn x male Cx. quinquefasciatus from Castelli, which did not happen (Table IV). An intracellular rickettsial agent of the genus Wolbachia found in reproductive tissues of arthropods (including Cx. pipiens) has been reported as having a profound effect on the host's population structure. If the sperm from a Wolbachia-infected male fertilizes an uninfected egg, unidirectional cytoplasmic incompatibility can occur, producing zygotic death. Different factors as bacterial strain, host genotype and bacterial density seems to influence strength and direction of that incompatibility (Werren 1997). In our case, Castelli and Puerto Madryn populations showed the highest genetic distance; however, 76.5 to 100% of hatching was recorded for interpopulation crosses. It is possible that the population from Rosario was infected by a Wolbachia strain not present in other populations tested. This may explain the important unidirectional reduction in fertility observed in some crosses between males from Rosario and females from genetically similar populations. The finding of hybrid individuals in samples from Cordoba and Rosario, and the fertility of the hybrids obtained under laboratory conditions (Brewer et al. 1987, Almiron et al. 1995), plus data on genetic distance and gene flow here presented, provide genetic evidence about the subspecific status of Cx. pipiens and Cx. quinquefasciatus in Argentina. The recognition of the existence of an important gene flow between intermediate populations of both forms provides useful information on their dispersal potential, and possibly of the infectious agent they transmit. This knowledge may also afford data for any attempt to perform genetic control on the species. Acknowledgment To Dr Antonio Blanco for the critical revision of the manuscript; to Dr Thomas P Monath, who kindly supplied materials used in this study, and Dr Esteban Bakos for providing us mosquitoes from Chaco Province. This work was supported, in part, by the Consejo Nacional de Investigaciones Cientificas y Tecnicas (CONICET), Argentina, and the Consejo de Investiga-ciones Cientificas y Tecnologicas de la Provincia de Cordoba (CONICOR). REFERENCES Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98011b.jpg] [oc98011e.jpg] [oc98011c.jpg] [oc98011a.jpg] [oc98011f.jpg] [oc98011d.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}