 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Genetic Variation among Natural and Laboratory Colony Populations of Lutzomyia longipalpis (Lutz & Neiva, 1912)(Diptera:Psychodidae) from Colombia Gregory C Lanzaro/^+, Bruce Alexander*^++, John-Paul Mutebi, James Montoya-Lerma*, Alon Warburg**
Department of Pathology, Center for Tropical Diseases, University of Texas
Medical Branch, Galveston, TX 77555-0609, USA Accepted 5 November 1997; Received 22 April 1997
Code Number:OC98013
Sizes of Files:
Text: 19.9K
Graphics: Line drawings and photographs (jpg) - 31.4K
Tables (jpg) - 74.3K
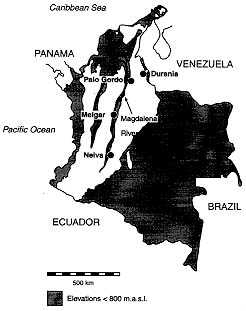
Genetic diversity among three field populations of Lutzomyia longipalpis in Colombia was studied using isozyme analysis. Study sites were as much as 598 km apart and included populations separated by the eastern Cordillera of the Andes. Genetic variability among populations, estimated by heterozygosity, was within values typical for insects in general (8.1%). Heterozygosity for field populations were compared with a laboratory colony from Colombia (Melgar colony) and were only slightly lower. These results suggest that establishment and long term maintenance of the Melgar colony has had little effect on the level of isozyme variability it carries. Genetic divergences between populations was evaluated using estimates of genetic distance. Genetic divergence among the three field populations was low (D=0.021), suggesting they represent local populations within a single species. Genetic distance between field populations and the Melgar colony was also low (D=0.016), suggesting that this colony population does not depart significantly from natural populations. Finally, comparisons were made between Colombian populations and colonies from Brazil and Costa Rica. Genetic distance values were high between Colombian and both Brazil and Costa Rica colony populations (D=0.199 and 0.098 respectively) providing additional support for our earlier report that populations from the three countries represent distinct species. Key words: sand fly - Lutzomyia longipalpis - population genetics - species complex The geographical distribution of Lutzomyia longipalpis sensu lato extends from central Mexico to northern Argentina and Paraguay (Young & Duncan 1994). This species is unusual among New World sand flies in its ability to occupy a variety of habitats, including those that have been considerably disturbed by man. Lu. longipalpis sensu lato occurs in close association with man and is the principal vector of Leishmania chagasi, causative organism of American visceral leishmaniasis. Mangabeira (1969) reported morphological differences among populations of Lu. longipalpis in Brazil. The differences he observed involved the number of spots on male abdominal tergites. Ward et al. (1983) carried out cross-mating experiments using laboratory-reared flies. They suggested that the reduced capacity of males to inseminate females of strains bearing a different number of tergal spots indicated that Lu. longipalpis is a complex of at least two sibling species. Lanzaro et al. (1993) analyzed genetic variability in laboratory colonies of Lu. longipalpis from Costa Rica, Colombia and Brazil at 27 enzyme coding loci. Values for genetic distance among these populations were high and cross-mating experiments resulted in the production of sexually sterile male progeny, suggesting that colony populations represent three species rather than one. In Colombia Lu. longipalpis occurs along the valley of the Magdalena River from the departments of Santander in the northeast to Huila in the southwest (Fig. 1). It is also present on the east side of the eastern Cordillera, a division of the Andes that rises to over 3000 m for much of its length, reaching a maximum altitude of 5493 m in the Sierra Nevada del Cocuy. The latter population may be contiguous with that found in the Venezuelan states of Merida and Trujillo. In the Colombian department of Norte de Santander Lu. longipalpis has been found in coffee plantations at 800 m above sea level (B Alexander, unpublished) and this together with the absence of cases of visceral leishmaniasis due to L. (L.) chagasi in the area suggests that this population may differ genetically from that of the Magdalena valley.
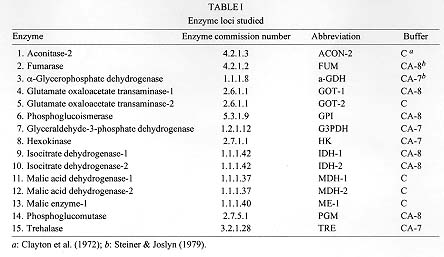
MATERIALS AND METHODS Sampling information - Adult sand flies were collected on June 4-5 1993 by aspirating them from human or animal bait in a stone-walled enclosure containing pigs adjoining a coffee plantation at Finca La Primavera, Morreton, near Durania, department of Norte de Santander; on June 7 1993 in pastureland near the district of Palo Gordo, between Giron and Piedecuesta, near the city of Buca-ramanga department of Santander; and on June 10 1993 in a scrub forest on the outskirts of Neiva, department of Huila (Fig. 1). Durania and Palo Gordo are 95 km apart, but are separated by the crest of the Eastern Cordillera of the Andes, which at this point rises to an elevation of over 3000 m. Durania lies on the eastern slope, at an elevation of 1000 m, Palo Gordo is an agricultural region in the Magdalena valley on the west slope, at an elevation of 500 m. Durania and Palo Gordo lie 598 and 500 km respectively northeast of Neiva. Neiva lies at an elevation of 577 m in the extreme southwest portion of the Magdalena valley. Insects were transported back to the laboratory in standard plaster-lined containers and either immobilized by freezing before being preserved dry in liquid nitrogen prior to analysis or placed in cages for the establishment of new laboratory colonies. In addition to the above mentioned field collected material, flies from the following three laboratory strains were analyzed for comparison: (1) the Brazil strain, which originated in Lapinha caves near Belo Horizonte, Minas Gerais (approximate coordinates 19 degrees 33'S, 44 degrees 0'W); (2) the Colombia colony derived from flies collected by RB Tesh in the village of El Callejon, near Melgar, Tolima department (4 degrees 11'N, 74 degrees 18'W) in 1989; (3) the Costa Rica colony was started with flies collected by us near Liberia, Guanacaste province (10 degrees 37'N, 85 degrees 26'W) in 1991. New colonies were established from Palo Gordo and Neiva. All colonies were maintained as described by Modi and Tesh (1983). Isozyme analysis - Field collected sand flies were identified to species by morphological characteristics and stored in liquid nitrogen. The whole bodies of individual adult flies were homogenized in 7 ml of distilled water. Homogenates were applied to horizontal 12.5% w/v starch gels for electrophoresis using standard procedures (Steiner & Joslyn 1979). Two different buffer systems were employed to maximize electrophoretic separation of enzymes. These are given with references in Table I. Enzyme-specific histochemical staining procedures were employed to visualize bands on gels. Discrete zones of staining activity on gels were assumed to be controlled by single loci coding for specific enzyme products. The products of 16 putative enzyme loci provided adequate resolution to study variability (Table I). Loci coding for the same enzyme were designated numerically in sequence from most cathodal to most anodal. Alleles were scored on the basis of the distance bands migrated through the gel with reference to the most common allele.
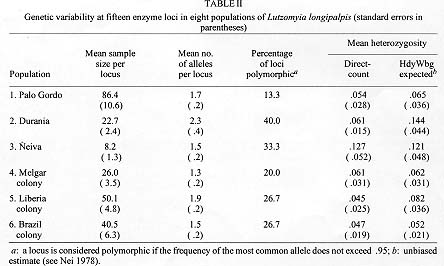
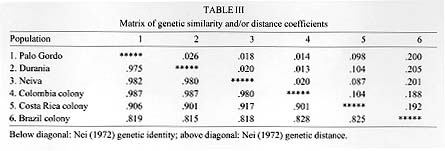
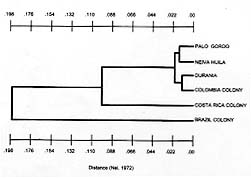
RESULTS Measures of genetic variability are presented in Table II. The Neiva population was most variable, with heterozygosity at about 13%, while the Palo Gordo population was most uniform (heterozygosity of 5.4%). The laboratory colony originating from near Melgar had comparable levels of genetic variability (heterozygosity=6.1%). Levels of genetic divergence and estimates of genetic relatedness between populations were made using genetic distance and similarity statistics (Table III). There were no diagnostic alleles. The average genetic distance among field populations (Durania, Palo Gordo and Neiva) was low, D = 0.021, suggesting these represent local populations within a single species. Comparison of these three populations with a sample from the Melgar laboratory colony gave a slightly lower average genetic distance of D = 0.016. These relationships are illustrated as a dendrogram (UPGMA) of genetic distances between laboratory and field populations (Fig. 2).
DISCUSSION Heterozygosity in three field populations (Fig. 1) of the Colombian sibling species of Lu. longipalpis was found to be 8.1%. This value is consistent with that published by Nevo (1978) in a survey of 23 insect species, suggesting that in terms of genetic variability the Colombian sibling species of Lu. longipalpis is a typical insect species. This value is, however, lower than the average heterozygosity (12.4%) published for Diptera (Graur 1985). Heterozygosity in the Melgar laboratory colony was found to be only slightly lower, 6.1%. Morrison et al. (1995) also compared heterozygosity in field populations of Lu. longipalpis from Colombia with the Melgar colony and although they report higher values, they also found little difference between field and colony populations (heterozygosity in five field populations = 14%, in the Melgar colony = 11%). Together these observations suggest that in the establishment of the Melgar colony the colonization process and maintenance in the laboratory over extended periods of time have resulted in very little effect on isozyme variability as estimated by heterozygosity. Levels of genetic divergence among populations was described using genetic distance (Nei 1972). Comparisons were made at three levels: between the three Colombian field populations; between Colombian field populations and the Melgar (Colombia) colony and between Colombian field populations and colony populations of different Lu. longipalpis sibling species (Brazil and Costa Rica). Three local populations were compared (Fig. 1). Values for genetic distance, D, among the three field populations was very low (D = 0.021), consistent with values among local populations of a single species. This suggests that the genetic composition of these populations is maintained by appreciable levels of gene flow (migration). Wright's FST statistics were calculated and migration rates (Nm) estimated using the relationship Nm = ¼ (1/ FST - 1). Migration rates were low, Nm = 2, but this value is high enough to prevent the evolution of significant levels of genetic divergence among populations. This value is appreciably lower than those reported by Morrison et al. (1995), but this is not surprising since the populations they studied were not more than 25 km apart, whereas our sites were separated by much greater distances (95-598 km) and by formidable physical barriers as well (the Andes Mountains). We compared the level of genetic divergence between three field populations and the Melgar laboratory colony and found little genetic divergence, D = 0.016 (0.013-0.020). These results depart from those of Morrison et al. (1995), who found a significantly higher genetic distance between field populations and the Melgar colony (D » 0.14). This led them to conclude that high levels of genetic change can be associated with the colonization process and to warn that analysis of collections directly from the field are essential before inferences about taxonomic relationships can be made. Our results suggest the contrary, that laboratory colonies of Lu. longipalpis can be developed which do not depart significantly from natural populations and may therefore accurately reflect genetic relationships among them. The final set of comparisons based on genetic distance involved relationships between populations of Colombian Lu. longipalpis species and sibling species from Brazil and Costa Rica. Genetic distance between Colombian populations and individuals from the Liberia Costa Rica colony was high, D = 0.098. Similarly, genetic distance between Colombian populations and the Lapinha Cave Brazil colony was also high, D = 0.199. Relationships among all populations are best visualized by examination of the dendrogram constructed using the UPGMA method and illustrated in Fig. 2. There are three distinct population groups apparent on the dendrogram: Colombia, which includes four populations only slightly differentiated from each other; Costa Rica; and Brazil. These results support earlier work (Lanzaro et al. 1993) that resulted in the description of Lu. longipalpis as a complex of at least three sibling species occurring in Colombia, Costa Rica and Brazil. In conclusion, the three populations studied here appear to represent local populations within a single species, despite their being separated by substantial distances and formidable geographic barriers. We found no significant genetic differentiation between samples collected directly from natural populations in Colombia and a long-standing laboratory colony originating from Colombia. Finally, in comparisons based on genetic distance, our results provide additional support for our earlier conclusion that Lu. longipalpis in Colombia represents a distinct species from that found in either Central America (Costa Rica) or Brazil. Acknowledgements To Javier Palta and Paulina Fajardo for assistance in handling specimens and maintaining laboratory stocks in Cali; to Victor Manuel Angulo and Yolanda Gamarra for assistance in making collections from Palo Gordo; to Scott Mariott for assistance in isozyme studies and maintenance of sand fly colonies in the laboratory in Texas. This research was supported by a grant from the John D. and Catherine T. MacArthur Foundation program entitled `Molecular Biology of Parasite Vectors' and from grant AI39540 from the National Institutes of Health to the first author and grants SP50 A1 30603 from the National Institutes of Health and 2229-04-004-92 from COLCIENCIAS Project I to Dr Nancy G Saravia, Executive Director, C.I.D.E.I.M. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98013b.jpg] [oc98013d.jpg] [oc98013c.jpg] [oc98013e.jpg] [oc98013a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}