 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
RESEARCH NOTE In vitro Method for Isolation of Amastigote Forms of Leishmania amazonensis from J774G8 Macrophage Induced by Temperature Shifting Maria Lucia Vieira Moreno, Maria de Nazareth Leal de Meirelles*/^+
Laboratorio de Patologia e Biologia Celular, Centro de Pesquisa Gonçalo
Muniz, Fiocruz, Salvador, BA, Brasil This work was supported by CNPq and Fiocruz. Received 9 January 1997; Accepted 24 September 1997
Code Number:OC98017
Sizes of Files:
Text: 13.7K
Graphics: Line drawings and photographs (jpg) - 63.4K
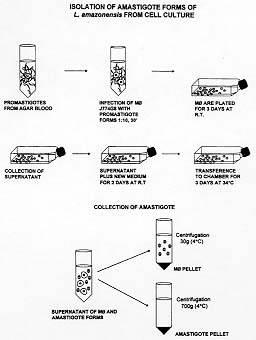
RESEARCH NOTE Key words: Leishmania - J774G8 macrophage - amastigotes - isolation - temperature shifting Leishmania parasites undergo a digenetic life cycle with the parasite migrating between flebotomus' midgut and mammalian host phagolysosomes. Most of the knowledge about biochemistry and molecular biology of the Leishmania parasite is related with the stage inside the insect vector that has been relatively easily cultured. The obtention of large amounts of intracellular amastigote stage which is responsible for disease in the mammalian host became an ultimate aim in order to perform biochemical, molecular and ultrastructural characterization. Several attempts have been done in order to establish an in vitro system to isolate the intracellular forms of the cutaneous leishmaniasis parasites from animals infections using cell-free media (DT Hart et al. 1981 Parasitology 82: 345-355, AA Pan 1984 Exp Parasitology 58: 72-80, PA Bates et al. 1992 Parasitology 105: 193-202, AA Pan et al. 1993 J Euk Microbiol 40: 213-223, AT Bijovsky 1994 Parasitol Res 8: 696-698, VH Hodgkinson et al. 1996 Exp Parasitol 83: 94-105) or from in vitro cultures of infected macrophages peritoneal exsudate or macrophage-like cell lines (E Handman & M Shapira 1977 Z Parasitenk 53: 75-81, KP Chang 1980 Science 209: 1240-1242, RL Berens & J Marr 1981 Parasitology 82: 345-355). Many of these works emphasize the role of temperature and pH in this stage transformation (reviewed in D Zilberstein & M Shapira 1994 Annu Rev Microbiol 48: 449-470). The exposure of Leishmania promastigotes to mammalian temperatures in axenic cultures induced morphological alterations similar to those occuring during the transformation of pro-mastigotes into amastigotes inside mammalian macrophages (S Eperon & D McMahon-Pratt 1989 J Protozool 36: 502-510). The combination of an elevation of temperature and a decrease in pH has produced stable axenic amastigote-like cultures (reviewed by P Bates 1993 Parasitol Today 9: 143-146). We developed an in vitro system for obtention of purified amastigote forms of L. amazonensis from monolayers of macrophages J774G8, originally derived from macrophages of the oil-induced peritoneal exudate of BALB/c mice (P Ralph et al. 1975 J Immunol 114: 898-900). Lesion amastigote forms were isolated and transformed to promastigote forms by weekly passages in a diphasic medium containing 4% (w/v) blood-agar base (DIFCO) and 15% (v/v) defibrinated rabbit blood at 25 C. The macrophage cell line J774G8 was maintained by serial passages at 37 C in Dulbbecco's modified Eagle medium (DME) supplemented with 10% inactivated fetal calf serum FCS, 2mM L-Glutamine and 100 uM gentamycin (pH 7.2). The maintenance of this cell line in culture is facilitated by the fact that these macrophages adhere loosely to the flask culture and can be washed off easily by vigorous agitation (Pan & Honigberg 1985 Z Parasitenkd 71: 3-13). By centrifugation, we recover the supernatant containing cells and put it to adhere in other flasks and the cells adhere and start to divide again. So, during the growh of this cell line, we have an adherent population and a released population in the supernatant. Our complete experimental assay included the following steps: (i) 5x10^6/ml of J774G8 cells obtained after centrifugation at 700 g for 10 min were ressuspended in DME and were infected with 50x10^6/ml stationary phase promastigote forms, at room temperature. After 30 min of parasite-cell interaction, the medium was transferred to a 30 ml flask and incubated at 25 C during 3 days (different sizes of the flask culture could be used depending of the inicial inoculum); (ii) after this period, the supernatant of the culture containing the J774G8 cell suspension and not interiorized parasites were collected and transferred with a new medium to two or more flasks and the cultures were incubated for two more days at 25 C; (iii) the next step of the assay was transfer the flasks to a temperature of 34 C with 5% CO2, for three days. At this time, we noticed that the pH of this medium was acidic around 6.5 and large amounts of amastigote forms were released from the macrophages; (iiii) collection of amastigotes forms were withdrawn by differential centrifugation including a 30 g centrifugation to separate the macrophages and a 700 g centrifugation to obtain the amastigote pellet. When promastigotes were engulfed by the J774G8 line, which does not mount an oxigen burst, the parasites were not killed (Titus et al. 1944 Immunol Ser 60: 437-459). Promastigotes need to transform to amastigotes within the cells before they start to multiply and there is also no lag period preceding growth and multiplication of amastigotes in macrophages (Pan & Honigberg 1985 Z Parasitenkd 71: 3-13). The transformation of Leishmania promastigote to amastigote forms during the infection of J774G8 macrophages must involve a series of steps to allow not only the morphological transformation but the development of the normal physiological processes at higher temperature and its survival within the parasitophorus vacuole. Morphological parasite transformation might be a process of temperature induced (PS Doyle et al. 1991 Exp Parasitol 73: 326-334). It has been described that Leishmania spp. contains a proton-translocating ATPase in the plasma membranes that is responsible to create an electron-chemical gradient of protons mediating the intracellular pH homeostasis and also regulating their intracellular pH (TA Glaser et al. 1992 Mol Biochem Parasitol 51: 9-16, D Zilberstein & DM Dwyer 1988 Biochem J 263: 13-21). A Ca^2+ ATPase was purified, characterized and found firmly bound to the plasma membrane vesicles in some Leishmania spp. (G Benain & PJ Romero 1990 Biochem Byophys Acta 1027: 79-84, J Ghosh et al. 1990 Biol Chem 265: 11345-11351). Recent reports using in situ ultrastructural detection showed the presence of a Ca^2+ ATPase on isolated parasites and also at the plasma membrane of the parasite when inside the phagosome of infected host cells. This finding points to a role of this enzyme in the regulation of calcium levels for amastigote forms within the parasitophorus vacuole (S Corte-Real et al. 1995 J Submicrosc Cytol Pathol 27: 359-366) which could increase the stability of the amastigote plasma membrane within a microenvironment with high calcium concentration allowing its survival and proliferation (NK Ganguly et al. 1991 J Pharm Pharmacol 43: 1400-142). We performed comparative studies with the amastigote forms obtained both from cell culture and from animal lesions and observed that they displayed different sizes. Amastigote forms collected from cell cultures were larger than those from animal lesions measuring 4.2 x 2.0 um and 3.3 x 1.5 um, respectively. The scheme representative of the experimental assay we develop for Leishmania amastigotes isolation from cell cultures is showed in the Fig. 1.
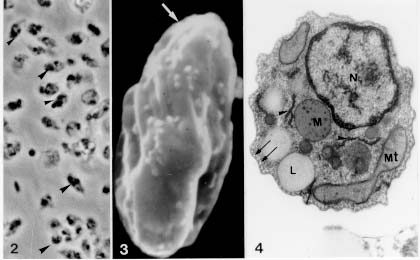
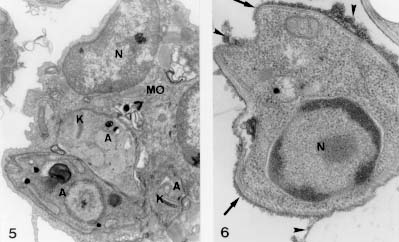
Figure 1 : schematic graphic showes the isolation of Leishmania amastigote forms from macrophage cell line J774G8 induced by temperature shifting. The following steps were followed: (i) infection of macrophages with promastigote forms of Leishmania for 30 min and platting them for three days at R.T.; (ii) collection of supernatant of the infected macrophage culture plus free parasites for incubation for two days at R.T; (iii) transference of the cell culture to a 34 C, for three days; (iiii) collection of the liberated amastigotes from the macrophage by differential centrifugation. Phase contrast microscopy of amastigote forms obtained as described displayed round forms without flagellum (Fig. 2). For scanning electron microscopy, after 2.5% gluraldehyde (GA) in cacodylate buffer fixation and 1% osmium tetroxide (OsO4) post-fixation the cells were critical point dryed. Amastigote forms displayed an oval shape, no extracellular flagellum and a wavy plasma membrane (Fig. 3). Figures 2-4: aspects of the suspension of the purified Leishmania amastigote forms shown by phase contrast microscopy. They displayed an oval shape without flagellum. 1.600 X. Fig. 3: scanning electron microscopy: amastigote forms displayed ondulations at the plasma membrane and no extracellular flagellum. 18.000 X. Fig. 4: transmission electron microscopy: amastigote forms showed presence of sub-pelicular microtubules near the plasma membrane (arrow), mitochondria (MT), lipid granules (L), endoplasmic reticulum profiles (arrowhead) and megasomes (M). 32.000 X. For transmission electron microscopy analysis, the cells were fixed for 1 hr with 2.5% GA in cacodylate buffer at 4 C , followed by post-fixation with 1% OsO4, for 1 hr, dehydration in graded series of acetone and embbeding with Epon resin. Thin sections were obtained in a Reichert ultramicrotome OmU3, double stained with lead citrate and uranyl acetate and examined in a Carl Zeiss EM-10C microscope. The ultrastructural observation of these amastigotes forms released in the medium showed the presence of sub-pelicular microtubules, mitochondria, lipid granules, cisternae of endoplasmic reticulum and defined megasomes (Fig. 4). The megasomes are the amastigote-specific lysosome-like organelles of L. mexicana and related species that contain high levels of proteinase activity. Several Leishmania parasites displaying a k-DNA (kinetoplast) as a bar-like shape, were observed within the J774G8 macrophage (Fig. 5) We confirmed that the surface charge of amastigote forms is negative with the binding of the probe cationized ferritin (CF) to the plasma membrane anionic sites and followed the binding of CF from 15 min to 5 hr. Within 3-5 hr of the incubation shedding processes occured, showing mobility of its surface components (Fig. 6). Figures 5 and 6 Leishmaniasis is one of the major protozoan disease in several regions of the world and its control is based in the search for vaccines and powerful chemotherapic agents. These measures must be directed to the amastigote forms, the infective forms of the mammalian hosts. Working with temperatures shifts from 25 to 34 C, within an acidic environment in a simple, easy method free of contaminations of host-cell debris, we obtained quantities of the parasites equivalent to 1 x 10^6/ml of purified amastigote forms. These forms were morphologically and physiologically tested with ultrastructural studies and fitted the requirements of the already characterized amastigote forms. Our results of isolating amastigote forms from monolayers of the macrophage-like cell line of J774G8 allows the posssibility to work with large quantities of the parasites to perform studies with the intracellular stage of the Leishmania species including protein synthesis, gene expression, ultrastructural alterations, mechanisms of drug action and the development of strategies for disease treatment and prevention. Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98017c.jpg] [oc98017b.jpg] [oc98017a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}