 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Experimental Infection of Calomys callosus (Rodentia, Cricetidae) by Toxoplasma gondii
Silvio Favoreto-Junior, Eloisa AV Ferro^*, Diogo Clemente,^ Deise AO Silva,
Laboratorio de Imunologia, Departamento de Patologia Received 29 April 1997; Accepted 23 July 1997
Code Number:OC98018
Sizes of Files:
Text: 21.5K
Graphics: Line drawings and photographs (jpg) - 25K
photograph (gif) - 55.9K
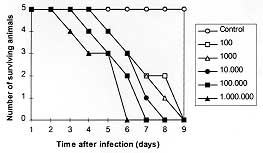
Calomys callosus, Rengger 1830 (Rodentia, Cricetidae), a wild rodent found in Central Brazil, was studied to investigate its susceptibility to Toxoplasma gondii experimental infection and its humoral immune response against this protozoa. The electrophoretic profile of the serum proteins of C. callosus showed that IgG, which shows no affinity to Protein A, has higher cross reactivity with rat IgG than with IgG from other rodents. The susceptibility assay was performed by inoculation groups of animals with various suspensions of T. gondii tachyzoites from 10^2 to 10^6 parasites. All animals died between 3 and 9 days after infection and the kinetics of antibody synthesis was determined. Basically, they recognized predominantly the immunodominant antigen SAG-1 (P30). The immunohistochemistry assays revealed that the liver was the most heavily infected organ, followed by the spleen, lungs, intestine, brain and kidneys. It can be concluded that C. callosus is an excellent experimental model for acute phase of Toxoplasma infection. Key words: Toxopasma gondii - Calomys callosus - experimental infection Toxoplasma gondii, an intracellular coccidian, infects a wide range of eukaryotic cells and it is an important opportunistic pathogen for humans and others animals (Jackson & Hutchison 1989). The infection is frequently assymptomatic but there are two groups of high-risk individuals: the human foetuses and the immunosuppressed patients, particularly those with acquired immunodeficiency syndrome (AIDS), that develop fatal toxoplasma meningoencephalitis. Several aspects of the mechanisms responsible for the pathological lesions leading to the clinical toxoplasmosis have been elucidated by using experimental models (Hutchison et al. 1982, Graham et al. 1984, Sims et al. 1988) and mouse has been the most employed animal in these studies (Gazzinelli et al. 1992). However, the experimental infection of wild rodents is poorly explored. On the other hand, it has been demonstrated that wild animal species present unique aspects when they are studied in investigation focusing host-parasite interactions (Deane et al. 1984, Jansen et al. 1985, Borges et al. 1992). Calomys callosus (Rodentia, Cricetidae) has been described as a reservoir for various infectious agents (Petter et al. 1967, Justines & Johnson 1970, Ribeiro 1973, Borges et al. 1992, Vaz-de-Lima et al. 1992). This rodent was also studied in its biology and breeding aspects, and since then this animal is being used in experiments with T. cruzi (Mello 1978, Mello et al. 1979). This paper presents the results of a study involving the experimental infection of C. callosus by T. gondii in order to evaluate the degree of its susceptibility to the parasite and its applicability as a model for experimental toxoplasmosis. MATERIALS AND METHODS Parasites - T. gondii parasites of the RH strain were maintained by serial passage in Swiss mice by standard procedure as described previously (Camargo et al. 1978, Mineo et al. 1980). Calomys callosus specimens - The specimens of C. callosus of the Canabrava strain used in this study belong to a resident colony housed at the Laboratory of Histology from Universidade Federal de Uberlandia and were kindly provided by Dr Judith Kloetzel. The animals were kept on a 12 hr light:12 hr dark cycle in a temperature controlled room (25 C) with food and water ad libitum. All animals were approximately 60 days old males, weighing about 25 g on inoculation. Serum samples were obtained from 20 animals and the immunoglobulins were isolated by precipitation on 40% ammonium sulphate solution. Both total serum proteins and immunoglobulins were analyzed by SDS-PAGE and submitted to western-blotting analysis after being transferred to nitrocellulose membranes (Burnette 1981). Experiments were carried out with protein-A and antibodies against rat, mouse, or rabbit in order to detect cross-reactivity to C. callosus immunoglobulins. Experimental infection - Six groups of five animals were used in these experiments. The animals of each group were inoculated intraperitoneally with 0.2 ml of various suspensions calculated to contain 10^2,^ 10^3, 10^4, 10^5 and 10^6 tachyzoites of T. gondii. Animals from the control group were inoculated with 0.2 ml media only. Parasites were obtained from mice peritoneal exsudate, washed twice in 1640 RPMI (Sigma Co., St. Louis, USA) containing 50 mg/l of gentamycin. The viable parasites were counted in Neubauer hemocytometer chamber in suspensions containing trypan-blue. Blood samples were collected from day 1 to day 9 after infection to determine the kinetics of antibody synthesis. Mortality was also observed up to nine days. As soon as the animals died, peritoneal exsudate was collected for parasite search. The next step was the collection of organs for histopathology and immunohistochemistry analysis. Liver, spleen, intestine, lung, testis, kidney, heart and brain were examined. ELISA for determination of anti-T. gondii antibodies in serum samples - An indirect immunoenzymatic test was employed as described (Camargo et al. 1978), with some modifications. Protein binding ELISA microtiter plates (Interlab, Brazil) were coated with 50 ul of a suspension of 1 x 10^6 tachyzoites of T. gondii/ml diluted in PBS. The plates were dried at 37 C and kept at -20 C until the serum samples were tested. The plates were then washed three times with phosphate-buffered saline containing 0.05% Tween 20 (PBS-T) and blocked with 5% low fat milk solution for 30 min at room temperature and incubated with the samples. Twofold dilutions of serum samples, diluted in PBS-T from 1:16 to 1:64, were added in duplicates to the plates and incubated for 45 min at 37 C. The third incubation step was carried out with the conjugate consisting of the Protein A-affinity purified rabbit IgG against C. callosus immunoglobulins labelled with peroxidase (type VI, Sigma, USA). After incubation with this conjugate for 45 min at 37 C, the plates were incubated with substrate solution consisting of H2O2 (Merck, Germany) and o-phenylenediamine (Merck, Germany) in 0.1M citrate-Na2HPO4 buffer (pH 5.5) for 15 min at room temperature. The reaction was stopped with 2N H2SO4 and the absorbance was read at 492 nm in a microwell reader system (Titertek Multiskan-Plus spectrophotometer, Flow Laboratories, USA). As controls, the serum samples and the conjugate were also incubated with uncoated plates in the same conditions. In addition, serum samples obtained from uninfected animals were also included. Immunohistochemistry for detection of parasitic antigens - For light microscopy, the material was dehydrated and embedded in paraffin. Sections of 3 um thickness were processed on glass slides. The preparations were hydrated by conventional techniques and incubated for 10 min at room temperature with 3% of H2O2 to block endogenous peroxidase. They were, then, treated with 2% of goat normal serum, diluted in phosphate buffered-saline (PBS), for 20 min at 37 C to block non-specific binding sites. The sections were incubated for 12 hr at 4 C with polyclonal antibodies from mice immunized with T. gondii antigens. Alternatively, the sections were incubated with mouse 6E9 monoclonal antibodies directed to SAG-1 (P30), the major surface antigen of the parasite obtained as described by Mineo and Kasper (1994). Controls to assess the binding specificity were run in parallel, with incubations without primary antibodies. Subsequently, the slides were rinsed in PBS and incubated with biotinylated goat anti-mouse IgG (Sigma Co., St. Louis, USA) for 30 min at 37 C. The signal of the reaction was amplified by ABC system (Vector Inc., Burlingame, USA) and revealed by 3'3'tetra-diamino-benzidine (DAB) (Sigma Co., St. Louis, USA) and counterstained by Meyer's hematoxiline. RESULTS The electrophoretic pattern of the immunoglobulins and the total seric proteins from C. callosus showed to be similar to other rodents as rat, rabbit and mice. The IgG molecule was detected as a 150 kDa band in the preparation obtained after fractionation of the serum proteins by ammonium sulphate. The western-blotting analysis showed that only antibodies reacting to rat IgG presented cross-reactivity with IgG from C. callosus. No affinity to protein A was observed to C. callosus IgG. All animals that were inoculated with T. gondii tachyzoites died at different times regarding the amount of parasites in the inoculum. No animal died in the control group. Results are shown in Fig. 1.
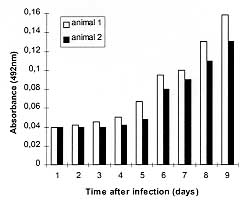
It was observed that two out of three survived animals at the group inoculated with 10^2 parasites presented antibodies to T. gondii between day 5 and 6, as detected by ELISA. The kinetics of this experiment is shown at Fig. 2. The western-blotting analysis demonstrated that these antibodies were reactive mainly against the 30 kDa epitope, the major surface antigen of T. gondii (data not shown).
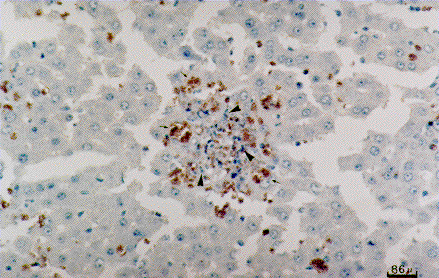
The parasitological examination revealed presence of tachyzoites in the peritoneal fluids of the animals as soon as they died. The immunohistochemistry assays demonstrated that parasites were present in all examined organs, except testis. The liver was the most intensively infected organ. Fig. 3 shows that the parasitophorous vacuoles are present in the cytoplasm of the hepatocytes which exhibited picnotic nucleus. It was also detected the presence of dividing parasites in the spleen, lung, intestine, brain and kidney, though the intensity of parasitism was lower in these organs when compared with the degree of parasitism in the liver (data not shown). This histopathological pattern was present in all examined animals and was not related with the number of inoculated parasites.
DISCUSSION The immune system components of C. callosus have been studied. Structural and functional similarities and differences with other organisms have been described. Homology between human and C. callosus complement components, particularly C3, C4 and B, has already been demonstrated (Vaz de Lima et al. 1992). The profile of hydrogen peroxide release from peritoneal macrophages from Trypanosoma cruzi infected mice and C. callosus shows some noteworthy differences, even though a parallel between macrophage activation and histopathological findings was observed in this two models (Borges et al. 1992). The present investigation showed cross-reactivity between IgG molecules from C. callosus and rat, demonstrating the existence of conserved epitopes which were not found among other studied rodents. Interestingly, C. callosus IgG shows no affinity to protein A which is one particular feature among other rodents. The data presented in this investigation demonstrated that C. callosus is highly susceptible to infection by RH strain of T. gondii. High mortality and massive presence of parasites in peritoneal cavity of the infected animals were found. Different susceptibilities to T. gondii infection were shown among various inbred and outbred laboratory rodents (Fujii et al. 1983, Zenner et al. 1993, Benedetto et al. 1996). On the basis of several criteria, the animals can be divided into three groups: (i) highly sensitive (S) as mice, Syrian hamsters and Chinese hamsters; (ii) moderately sensitive (M) as Mongolian gerbils and mastomyses ; and (iii) resistant (R) as rats and Korean striped field mice. S and M hosts supported intraperitoneal growth of the parasites and died of acute infection. M hosts showed prolonged time to death and low fatality as compared with S hosts. Some R hosts died only when they had received an extremely large size of inoculum, accompanying poor intraperitoneal proliferation of the parasite. Thus, our results demonstrated that C. callosus can be classified as highly sensitive host to T. gondii. The ELISA showed that two of three survived animals seroconverted between day 5 and 6 after infection. The antibodies were reactive mainly against the 30 kDa epitope, the major surface antigen of T. gondii, which is an important attachment factor to the parasites. It has been demonstrated that 30 kDa epitope is an immunodominant antigen in naturally infected hosts or in experimental toxoplasmosis. The antibodies reacting to this epitope belong to IgM and IgA isotypes in acute phase of murine toxoplasmosis (Mineo et al. 1993). In the present study, it was not possible to identify the involved isotype because the labelled secondary antibody showed reactivty to total C. callosus immunoglobulins. Jamra and Vieira (1991), studying the acute phase of toxoplasmosis in mice, observed the presence of parasites at peritoneal cavity and in various organs, with predilection to lung, followed by skeletal muscle, heart and brain. In our model, the results obtained by immunohistochemistry demonstrated that the liver from C. callosus was the most heavily infected organ, followed by spleen, lung, intestine, brain and kidney. These variances may be related to differences of the host or of the technique employed for parasite detection. According to our observations, C. callosus is proposed as an alternative model to study acute phase of experimental toxoplasmosis. The susceptibility capacity found to this host is important because it becomes useful feature for studying infection and immunity to toxoplasmosis. Indeed, we are now studying C. callosus as a model for congenital toxoplasmosis by looking on the invasion capacity of trophoblastic cells at in vivo experiments. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98018a.jpg] [oc98018b.jpg]Line drawing images[oc98018c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}