 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Identification of Novel Membrane Structures in Plasmodium falciparum Infected Erythrocytes Carlos A Clavijo, Carlos A Mora, Enrique Winograd^*/+
Laboratorio de Biologia Celular, Instituto Nacional de Salud, Avenida El
Dorado con Carrera 50, Bogota, Colombia Received 18 December 1996; Accepted 20 August 1997
Code Number:OC98022
Sizes of Files:
Text: 23.6K
Graphics: Line drawings and photographs (jpg) - 45.5K
Little is known about the molecular mechanisms underlying the release of merozoites from malaria infected erythrocytes. In this study membranous structures present in the culture medium at the time of merozoite release have been characterized. Biochemical and ultrastructural evidence indicate that membranous structures consist of the infected erythrocyte membrane, the parasitophorous vacuolar membrane and a residual body containing electron dense material. These are subcellular compartments expected in a structure that arises as a consequence of merozoite release from the infected cell. Ultrastructural studies show that a novel structure extends from the former parasite compartment to the surface membrane. Since these membrane modifications are detected only after merozoites have been released from the infected erythrocyte, it is proposed that they might play a role in the release of merozoites from the host cell. Key words: Plasmodium falciparum - parasite release - membrane fusion - parasitophorous vacuolar membrane The erythrocytic life cycle of the human malarial parasite Plasmodium falciparum is responsible for most of the pathology and mortality associated with this disease (Miller et al. 1994). The cycle is initiated by entry of a merozoite into the host red blood cell by invagination of the erythrocyte plasma membrane (Ward et al. 1993, Dluzewski et al. 1995). During the next 48 hr, the intracellular parasite develops surrounded by two membranes: the erythrocyte plasma membrane and the invaginated membrane closely apposed to the parasite itself, the parasitophorous vacuolar membrane. During the first 20 hr of development, the young feeding parasite (trophozoite) is observed as a "ring form". After this time, an increase in various metabolic activities including the degradation of hemoglobin in digestive vacuoles takes place. Digestion of hemoglobin results in the production of amino acids and an insoluble pigment called hemozoin, which accumulates in and is characteristic of the mature trophozoite stage. During the last 14 hr of development, DNA replication and cellular differentiation of 16 to 32 daughter cells or merozoites occurs by asexual division or schizogony. Hemozoin concentrates within a residual body, before merozoites are released from the red cell into the bloodstream. Little is known about the molecular mechanisms underlying the release of merozoites from malaria infected erythrocytes. Video-microscopy studies^ have shown that the actual rupture of a schizont-infected erythrocyte is preceded by swelling and vesiculation of the host cell membrane, and then merozoites are released with explosive suddenness (Dvorak et al. 1975, Hermentin & Enders 1984). Malarial proteases have been implicated in merozoite release, since rupture of infected erythrocytes can be prevented through the use of protease inhibitors (Banyal et al. 1981, Hadley et al. 1983, Lyon & Haynes 1986). Furthermore, plasmodial proteases with activities against known red cell membrane skeletal proteins, have been isolated and characterized (Deguercy et al. 1990); however it has not been determined whether these proteases actually participate in the rupture of the red cell membrane during merozoite escape. At the time when schizogony is nearly complete, the energetic charge of the infected erythrocyte falls (Yamada & Sherman 1980). Such a reduction in ATP levels could disturb the osmotic gradient across the erythrocytic membrane that is required for maintaining cellular volume. Possibly, pore forming proteins which have been reported in other parasitic protozoa (Noronha et al. 1994, Andrews 1994), could function in the escape of merozoites from the infected cell, but these are yet to be identified in the case of malarial parasites. In this study, a preliminary characterization of membranous structures found in the culture medium at the time when merozites are released from infected erythrocytes has been carried out. It is shown that membranous structures are composed of a surface membrane, a parasitophorous vacuolar membrane, a residual body containing electron-dense material, and a novel structure which extends from the former parasite compartment to the surface membrane. Our results suggest a possible function of these novel membrane modifications in the process of parasite escape from the host cell. MATERIALS AND METHODS Isolation of membranous structures - P. falciparum cultures (Trager & Jensen 1976) were synchronized by gelatin flotation (Jensen 1978) followed by sorbitol lysis (Lambros & Vandenberg 1979) of mature infected erythrocytes. Parasite maturation was monitored every 8 hr by Giemsa staining of thin blood-smears. When parasites reached the ring stage, the culture was centrifuged at 230 x g for 5 min. The supernatant containing membranous structures was centrifuged at 900 x g for 10 min, and the resulting pellet was resuspended in a small volume of 10 mM sodium phosphate buffer pH 7.4 containing 0.145 mM NaCl (PBS). The suspension of membranous structures was overlaid on top of a discontinuous gradient composed of 5 and 13% Percoll cushions (vol/vol in PBS), and centrifugation at 900 x g for 15 min was carried out. Membranous structures were isolated from the 5-13% Percoll interphase and washed three times in PBS. Fluorescence microscopy - Freshly isolated membranous structures were processed for immunofluorescence microscopy as previously described (Winograd & Sherman 1989). Rabbit anti-erythroid spectrin was kindly provided by Dr M Wasserman, and monoclonal antibody 8E7/55 against the parasitophorous vacuolar antigen QF116 was a kind gift from Dr AR Hibbs. Staining with merocyanine 540 was essentially carried out as previously reported (Kara et al. 1988). Membranous structures were labeled with quinacrine by incubation in a 0.1 % (w/v) quinacrine solution in Hepes Buffered Saline (HBS; 10 mM Hepes buffer pH = 7.40 containing 0,15 M NaCl) for 10 min at 37 C. After washing in HBS, cells were examined by fluorescence microscopy. As a control, live infected erythrocytes were labeled with quinacrine at different stages of development. In every instance, infected erythrocytes presented a strong fluorescence signal. Erythrocytes were surface labeled with biotin (Simpson et al. 1981) and infected with P. falciparum as described above. Membranous structures were isolated when parasites reached the ring stage of development. Membrane-bound biotin was detected by fluorescence microscopy after incubation in streptavidin-fluorescein. Electron microscopy - Membranous structures were purified by discontinuous Percoll gradients as described above, and immediately fixed for 1 hr in 2% glutaraldehyde in PBS at room temperature. Membranous structures were then processed either for transmission (Winograd & Sherman 1989) or scanning electron microscopy (SEM) (Gruenberg et al. 1983) and observations were either carried out on a Phillips transmission electron microscope Model CM10 or a Phillips scanning electron microscope Model 515. RESULTS To learn more about the process of merozoite release from malaria infected erythrocytes, we carried out a characterization of the membranous structures found in the culture medium of P. falciparum. The number of membranous structures in the culture medium increased every 48 hr (not shown), coincident with the time of merozoite release ( i.e. the temporal interphase between the schizont and ring stages), suggesting that these structures are implicated in the process of merozoite release from the infected erythrocyte. Observations by phase contrast microscopy, showed that membranous structures have morphology and dimensions similar to that of an infected erythrocyte, including the presence of a residual body containing dense material (Fig. 1A, C, E). By immunofluorescence microscopy, the outermost membrane reacted with antibodies directed against erythroid spectrin (Fig. 1B). Furthermore, when the erythrocyte membrane was labeled with biotin prior to infection, streptavidin conjugated fluorescein bound only to the outermost membrane of the structures (results not shown). These results suggest that the external membrane of the membranous structures originates from the infected erythrocyte.
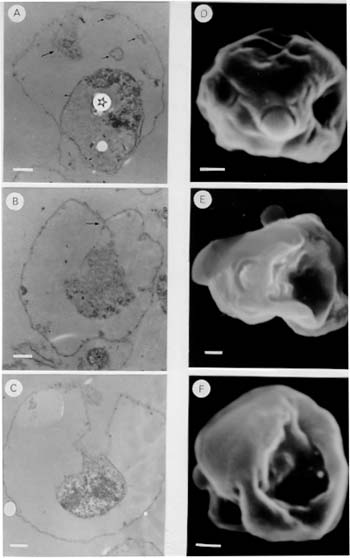
Membranous structures could be labeled with merocyanine 540 (Fig. 1D), a reagent previously shown to interact with the parasitophorous vacuolar membrane (Elford et al. 1985). In addition, membranous structures reacted with the monoclonal antibody 8E7/55 (Fig. 1F), whose specificity has been demonstrated to be against the parasitophorous vacuolar antigen QF116 (Kara et al. 1988). These results indicate that the parasitophorous vacuolar membrane is also an important component of membranous structures. When membranous structures were incubated with quinacrine no fluorescent signal was detected, suggesting that membranous structures are devoid of DNA. Based on the evidence presented above, membranous structures contain subcellular compartments expected to be present in a structure that arises as a consequence of merozoite release from the infected cell, i.e. it is composed of compartments not directly involved in the differentiation of merozoites (i.e. red cell membrane, parasito-phorous vacuolar membrane and residual body containing dense material). Observations using transmission electron microscopy, showed that membranous structures are composed of a surface membrane with associated knobs or excrescences and a residual body containing electron-dense material ( Fig. 2A). Vesicles or tubules, similar to membrane-bound structures described in schizont-infected erythrocytes (Langreth et al. 1978, Atkinson & Aikawa 1990), were found within the space defined by the outermost membrane and the residual body (Fig. 2A). In addition, larger membrane-bound structures (0.7 to 1.4 um) appearing to derive from the surface membrane, contain electron-dense material similar to the one present in the residual body (Fig. 2A). In other microscopic sections, the red cell membrane and the membrane surrounding the residual body appear to be continuous (Fig. 2B), and occasionally a well defined structure extending from the erythrocyte membrane to the residual body was observed (Fig. 2C). These results indicate that the former parasite compartment and the extracellular environment are communicated through a novel duct-like structure.
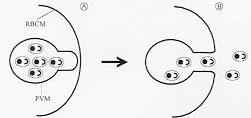
Scanning electron microscopic observations of the membranous structures, showed the presence of an opening which caves in appearing to lead to a duct-like structure (Fig. 2D, E, F). DISCUSSION In this study, membranous structures were identified in the culture medium of P. falciparum infected erythrocytes. The time of appearance of these membranous structures was found to be correlated to the event when merozoites are released from infected erythrocytes. Membranous structures were found to be composed primarily of the erythrocyte membrane, the parasitophorous vacuolar membrane, and a residual body containing dense material. Since these subcellular compartments do not participate in the cellular differentiation of merozoites, membranous structures could correspond to cellular structures arising periodically in the culture medium as a consequence of merozoite release from infected erythrocytes. Ultrastructural studies demonstrated the presence of a membrane extending from the former parasite compartment to the surface membrane, forming an apparent duct-like structure. The fact that these structures are not detectable on schizont-infected erythrocytes either by transmission (Langreth et al. 1978, Atkinson & Aikawa 1990) or SEM (Gruenberg et al. 1983), suggests that the formation of such membrane modifications might be functionally related to the release of merozoites from infected erythrocytes. In a previous study a parasitophorous duct on erythrocytes infected with mature stages of the parasite, through which macromolecules present in the external culture medium could reach the intraerythrocytic parasite, was proposed (Pouvelle et al. 1991). More recently, (Hibbs et al. 1997) evidence for a functional duct was not found. Although the exact role of the parasitophorous duct is not currently understood, we would like to suggest that the duct identified in this study, might play a role in the release of merozoites from infected erythrocytes (Fig. 3). The model of parasite release assumes that the parasite compartment within the infected cell becomes topologically continuous with the extracellular environment as a result of a membrane fusion event. Figure 3: a model of parasite release in malaria infected erythrocytes. A schizont infected erythrocyte is shown in (A). Upon membrane fusion between the parasitophorous vacuolar membrane (PVM) and the red blood cell membrane (RBCM), merozoites are released (not necessarily with the same orientation as depicted in the diagram) from the infected cell to the extracellular environment (B). It is proposed that membrane fusion occurs at sites where the parasitophorous vacuolar membrane and the erythrocyte plasma membrane are in close proximity. Such places could be regions where vesicles and large tubules derived from the parasitophorous vacuolar membrane extend to the erythrocyte membrane in the form of a membranous network (Helmendorf & Haldar 1993, Elford & Ferguson 1993). Since protease inhibitors are known to interfere with the release of parasites (Banyal et al. 1981, Hadley et al. 1983; Lyon & Haynes 1986), the fusion event might be preceded by cleavage of selected red cell membrane-skeleton components. The fusion of the two membranes would result in the formation of a duct-like structure whose dimensions permit merozoites to move out of the host cell by random diffusion. This model of merozoite release in P. falciparum infected erythrocytes would implicate an elaborated cellular machinery, where merozoite release and the formation of a duct-like structure could be highly regulated and coordinated to specific events of the parasite cell cycle. Studying the molecular components of this machinery could define new strategies for controlling infections in individuals afflicted with malaria disease. These findings might provide a conceptual framework to investigate whether evolutionary related organisms also have evolved similar mechanisms for parasite escape. Acknowledgments To Dr Moises Wasserman for allowing us to carry out the initial observations in his laboratory. To Dr Daniel Branton, Dr Vladimir Corredor, and Dr Irwin W Sherman for critically reviewing the manuscript. This investigation received financial support from the UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR), the Colombian Institute for the Development of Science and Technology "Francisco Jose de Caldas" COLCIENCIAS (research grant 22104-05-044-94), and Instituto Nacional de Salud. REFERENCES Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98022c.jpg] [oc98022b.jpg] [oc98022a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}