 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Succession and Abundance of Staphylinidae in Cattle Dung in Uberlandia, Brazil Jorge Anderson Guimaraes, Julio Mendes^+
Departamento de Patologia, Centro de Ciencias Biomedicas, Universidade
Federal de Uberlandia, Caixa Postal 593, 38400-903 Uberlandia, MG, Brasil
Received 24 February 1997; Accepted 12 May 1997
Code Number:OC98024
Sizes of Files:
Text: 18.4K
Graphics: Line drawings and photographs (jpg) - 26.4K
Tables (jpg) - 51.9K
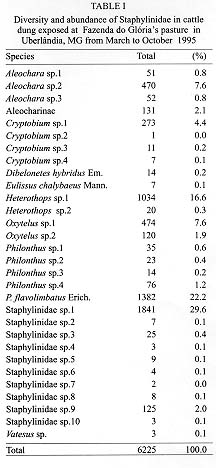
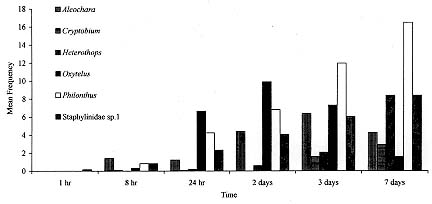
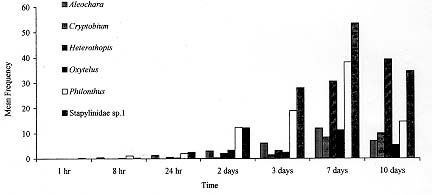
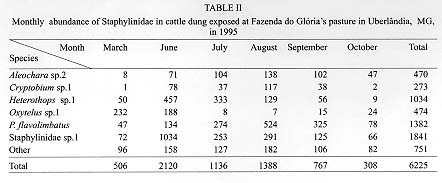
Fimicolous Staphylinidae prey on rearing dipterous in cattle dung, acting as their natural controllers, including pests such as horn fly. To survey the abundance and succession of these coleopterans in cattle dung deposited in pasture, six experiments were conducted from March to October 1995 in Uberlandia, State of Minas Gerais, Brazil. Cattle dung pats were exposed at a pasture for 1 hr, 8 hr, 24 hr, 2, 3, 7 and 10 days and were than taken to laboratory separate from each other, for Staphylinidae extraction. A total of 156 dung pats were exposed at pasture, from which 6225 Staphylinidae were recovered. Representing at least 30 species. Staphylinidae sp.1 (29.6%), Philonthus flavolimbatus (22.2%), Heterothops sp.1 (16.6%), Oxytelus sp.2 (7.6%), Aleochara sp.2 (7.6%) and Criptobium sp.1 (4.4%) were the most abundant, representing 87.8% from the total. The increased frequency of the majority of these species along the dung exposition time at pasture, indicated tha, they would be preying on at all the immature stages of the dipterous, or eggs and first instar larvae of species that lay eggs on the dung after its second exposition day at the pasture. Key words: Staphylinidae succession - cattle dung - horn fly - biocontrol - Brazil Cattle dung is a microhabitat for an abundant and diverse arthropod fauna (Blume 1985). Diptera, Coleoptera and Hymenoptera are the most abundant arthropods associated with dung pats in pastures (Poorbaugh et al. 1968, Nibaruta 1982, Blume 1985, Cervenka & Moon 1991, Mendes 1996). A portion of the fauna use dung pats as a breeding medium while the other part visit them to parasite or to prey on the coprophagous fauna. The most important biotic mortality factors of the dung-breeding Diptera are competitors dung-burying scarabs, the Staphylinidae and Histeridae predators (Coleoptera), and the parasitoids (Hymenoptera) (Wingo et al. 1974, MacQueen & Beirne 1975, Roth et al. 1983, Fay et al. 1990). The dung-breeding flies are responsible in part for cattle dung decay. On the other hand, when adults, the horn fly Haematobia irritans (Linnaeus 1758), the face fly Musca autumnalis (De Geer 1776) and the bush fly Musca vetustissima Walker 1857 (Diptera: Muscidae) are important pests of cattle and can cause expensive economic losses (Drummond et al. 1981, Lancaster & Meisch 1986, Schreiber et al. 1987). The horn fly is a specific blood-sucker of cattle hosts. The fly remains on the host's skin piercing constantly and leaves it only to deposit its eggs on fresh cattle droppings in pastures or to pass to another host (MacLintock & Depner 1954). This fly was recently registered in the North of Brazil (Valerio & Guimaraes 1983). Today it is seen infesting cattle in almost all of the country as well as the other neighboring countries of the American South Cone (Campano & Avalos 1994). Some arthropods visit or breed in cattle dung concomitantly with immature horn fly. Some of them are their natural enemies. Many researches in the North America, South Africa and Australia have reported the Staphylinidae species diversity in cattle dung and their importance as natural controllers of horn fly and face fly (Wingo et al. 1974, Harris & Oliver 1979, Roth et al. 1983, 1988, Doube 1986, Davis et al. 1988, Doube et al. 1988, Hunter et al. 1989, Fay et al. 1990). Information relating to the period of occurrence and abundance during dung decay are some of the essential basic knowledge about the pest enemies to be used in biological and or integrated control of dung-breeding pests (Wingo et al. 1974, Hunter et al. 1986). With the exception of the researches of Flechtman et al. (1995) and Mendes (1996) the fimicolous Staphylinidae are poorly investigated in Brazil. Therefore, the goal of this work was to survey the abundance and succession of Staphylinidae species during the first 10 days of cattle dung pat decay at a pasture in Uberlandia, State of Minas Gerais, Brazil. MATERIALS AND METHODS The study area was a Paspalum notatum Flugge pasture at Fazenda do Gloria which belongs to the Universidade Federal de Uberlandia (UFU). It is located about 5 km from Uberlandia, (18 degrees 55'S, 48 degrees 17'W). Dung pats from Holstein dairy cows were collected early in the morning after defecation on a cemented floor at the farm stables. The dung pats were placed in plastic pails, mixed up into dung pats and exposed as such at the pasture. During the months of June, July and August 1995, 24 circular dung pats, around 30 cm in diameter and weighting each about 1 kg, were laid 5 m apart on the experimental pasture. For March, September and October of the same year the number of the dung pats exposed in the same experimental pasture yard was 28. After 1 hr, 8 hr, 24 hr, 2, 3, 7 and 10 days (in June, July and August = the cooler months) 4 dung pats were collected with about 5 cm of the underlying soil and taken, separate from each other, to the UFU's laboratory of parasitology. Thereafter, two of the four dung pats and its associated soil were transferred to individual Berlese Funnels each of which was equipped with a 60W light bulb and these were kept for 5 days or until the funnel's content was conspicuously dry. This technique was used for the extraction of living Staphylinidae beetles from the dung and soil samples described above. The Staphylinidae extraction from the other two pats was hand made using tongs. All the specimens obtained were preserved in 70% alcohol. The climatic data were obtained at 5^o Meteorological District from Uberlandia, located about 6 km from the experimental area. To test for differences among the Staphylinidae abundance and the exposition time of the dung pats at the pasture a one way ANOVA analysis was performed. The means for each main factor and for each species were compared using the multiple comparisons F- test of Tukey. To test for the relationship between the abundance of the species or genera and the respective monthly mean temperatures of the performed experiments Spearman's Correlation Analysis was also performed. All the tests were performed using the Basic Statistics and Tables and Nonparametric Statistics & Distribution Fitting procedures of STATISTICA 4.5 for Windows (StatSoft, Inc. 1993). RESULTS The rain precipitation during the months of March, September and October were 239.1 mm, 22.0 mm and 65.2 mm and the respective mean temperatures were 23.9 C, 23.3 C and 24.0 C. The months of June, July and August presented respective climatic data: 3.4 mm, 1.6 mm and 0.0 mm and 19.1 C, 20.5 C and 22.6 C, being apparently drier and cooler than the other three months. At the experiment's end 156 dung pats had been exposed in the pasture yard area and a total of 6225 adult Staphylinidae belonging to at least 30 species had been collected (Table I). Staphylinidae sp.1, Philonthus flavolimbatus, Heterothops sp.1, Oxytelus sp.2, Aleochara sp.2 and Criptobium sp.1 were distinguished from the other species by their abundance, representing 87.8% of the total (Table I). Nevertheless, the relative frequency of these groups changed between experiments and with dung age (Table I, Fig. 1, Fig.2). The majority of the Staphylinidae presented increased abundance along the dung exposition time at the pasture. The abundance peak occurred in the third day of the dung pats and beyond (F = 17.64; p < 0.001) (Figs 1, 2). The exception was the Oxytelus sp.1 in the warmer months, for which the peak of abundance was recorded in the second and third days of the dung exposition. The majority of the most abundant species showed considerable inverse correlation with the monthly mean temperature (-0.48 < r <- 0.3; p < 0.001). They were more abundant in the drier and cooler months and even approached the data from the time exceeding ten days of the dung age (Table II). The exceptions were Oxytelus sp.1 which did not show correlation with temperature and Aleochara sp.2 that presented direct correlationed with this abiotic factor (r = 0.38; p < 0.001).
Figure 1: Staphylinidae succession in cattle dung pats exposed at Fazenda do Gloria pasture in Uberlandia in March, September and October 1995. Showed values are mean frequency by dung pat in each exposition time. Figure 2: Staphylinidae succession in cattle dung pats exposed at Fazenda do Gloria pasture in Uberlandia from June to August 1995. Showed values are mean frequency by dung pat in each exposition time. It was expected that the species would be most abundant in the warm months because of the great abundance of prey in the dung in this period (Laurence 1955, MacQueen & Beirne 1975, Nibaruta 1982, Doube 1986). In spite of the hight Staphylinidae abundance occurring in cool and the dry months, it is important to point out that the data are not sufficient for good inferences about their seasonallity, this will possibly be clarified with an increase in the number of experiments and a more representative number of months. The climatic changes did not apper to interfere significantly with the arriving time of the Staphylinidae at the dung pats (Figs 1, 2). The arrival of Aleochara spp., Heterothops spp., P. flavolimbatus and Staphylinidae sp.1 arrival at the dung pat within the first 24 hr after its exposition is indicative of their predatory tendency upon dipterous eggs and first instar larvae. On the other hand, the observed increase of the frequency could be due to the possible predation on other larval stages, on pupae and or on eggs and first instar larvae of some Diptera which layed eggs on the dung after the 2nd exposition day. The Staphylinidae access to such larvae and pupae would be enhanced by the dung tunnels previously made by larval Diptera and mature Scarabaeidae (MacQueen & Beirne 1975). The arrival of Oxytelus spp. at the dung pad in the first 24 hr of exposition probably indicates that it aimed to use it as food (Hanski 1987). While the Oxytelus spp. descending curve, between the third and seventh dung exposition days, could be due to the case of inadequacy of dung as food for such beetles. In the Aleochara spp. case, the increasing frequency would be related to predation along the first days and include egglaying in the later ones, since they are parasitoids when immature. The P. flavolimbatus frequency in two days old dung, were mostly below five individuals as pointed out by Harris and Oliver (1979), which responds to a significant reduction in H. irritans emergence. Nevertheless, there are many other species which are considered H. irritans natural enemies, as the cited above, occurr concomitantly with susceptible H. irritans immature stages in the dung. As in other regions, the Staphylinidae seem to be important natural controllers of Diptera in cattle dung in Brazil. The next step (apart from obtain more data about their seasonallity) would be to estimate predation efficiency data of the most abundant species on H. irritans. This study highlights Aleochara sp.2, Cryptobium sp.1, Heterothops sp.1, P. flavolimbatus and Staphylinidae sp.1 for such testing, corroborating Mendes' (1996) observations in the State of Sao Paulo. Acknowledgments To Drs Cecilia Lomonaco and Uriel Franco Rocha for the revision of the manuscript. REFERENCES Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98024d.jpg] [oc98024a.jpg] [oc98024c.jpg] [oc98024b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}