 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Life Cycle and Fecundity Analysis of Lutzomyia shannoni (Dyar) (Diptera: Psychodidae) Cristina Ferro/^+, Estrella Cardenas, Dario Corredor*, Alberto Morales, Leonard E Munstermann**
Instituto Nacional de Salud, Laboratorio de Entomologia, Avenida Eldorado,
Carrera 50, Santa Fe de Bogota, Colombia Received 18 June 1997; Accepted 11 December 1997
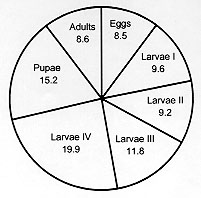
Code Number:OC98036 The life cycle of Lutzomyia shannoni (Dyar), was described for laboratory conditions with maximum daily temperatures of 27-30 C, minimum daily temperatures of 22-27 C and relative humidity between 87-99 %. Life cycle in each stage was as follows: egg 6-12 days (ave. 8.5 days); first stage larva 5-13 days (ave. 9.6 days); second stage larva 4-13 days (ave. 9.2 days ); third stage larva 5-19 days (ave. 11.8 days); fourth stage larva 7-37 days (ave. 19.9 days); pupa 7-32 days (ave. 15.2 days). The life expectancy of adults ranged from 4 to 15 days (ave. 8.6 days). The entire egg to adult period ranged from 36 to 74 days (ave. 54.6 days). On average, each female oviposited 22.7 eggs; the average egg retention per female was 24.3 eggs. Key words: Lutzomyia shannoni - forest sand flies - life cycle - fecundity - larval description - Colombia The sand fly Lutzomyia shannoni (Dyar) has a broad geographical distribution in Western Hemisphere from northern Argentina to southeastern United States. However, its distribution is highly disjunct within that range, depending on locally occurring environmental factors such as frequency of precipitation, temperature, physical barriers, habitat availability, and the distribution and abundance of vertebrate hosts (Young & Arias 1992). Lu. shannoni was first recorded in Colombia in 1966 in tree holes at the Chigorodo location, Departament of Antioquia (Osorno et al. 1972). Since then it has been recovered from Shannon traps and resting sites on large trees from sea level up to approximately 1000 m (Young 1979). In the United States of America Lu. shannoni is an important vector of vesicular stomatitis virus (VSV), a disease of cattle and sheep. Comer et al. (1991, 1994) recorded Lu. shannoni as the only sand fly present in the Ossabaw Island in Georgia and verified that this is the insect vector of VSV there. In Panama and Colombia specimens of Lu. shannoni have been found infected with unidentified flagelates (Johnson et al. 1963, Morales et al. 1981, Young & Lawyer 1987, Alexander et al. 1992). Althougth Lu. shannoni has not been associated with the transmission of severe disease in this country, it has been collected in the peridomestic environment (Pardo et al. 1996), inside houses and during human landing-biting captures (Alexander et al. 1992). Although Lu. shannoni has been bred in the laboratory for several generations (Young et al. 1981, Endris et al. 1982) and experimentally infected with Leishmania mexicana (Lawyer et al. 1987), L. panamensis (unpublished data from the Laboratory of Entomology, Instituto Nacional de Salud) by feeding on infected hamster, and with L. chagasi (Endris et al. 1982) through membranes, very little is known about its life cycle, developmental times, mating, oviposition and larval nutrition. Although effects of colonization may strongly affect genetic constitution and behavior of a species (Munstermann 1994, Morrison et al. 1995), life table parameters for many insect species of medical importance cannot be obtained in the field. For the past five years, the Entomology Laboratory of the Colombian INS has maintained a colony of Lu. shannoni that was initiated from specimens collected in October, 1992 in Carmen de Bolivar (Departament of Bolivar). The objective of the study is to provide baseline data concerning the life cycle and fecundity of this colony. MATERIALS AND METHODS The study began at the twenty-second generation of the Lu. shannoni colony. The laboratory conditions during the study were as follows: relative humidity between 80 and 97% (ave. 88%), maximum temperature between 27 and 30 C (ave. 28 C), and minimum temperature between 22 and 27 C (ave. 25 C). The adults were kept in the same rearing chambers as used for immature forms (Fig. 1). Males and females were passed to a cloth cage only for blood feeding. The rest of the procedure was done according to Endris et al. (1982) and Modi and Tesh (1983). The females were fed on hamster blood two days after emergence. For blood feeding the females, approximately equal numbers of males and females were transferred from the rearing chamber (Fig. 1) to cloth cages (20 cm x 20 cm). Mating was observed in these cages and the rearing chambers. An anesthetized (sodium pentothal, 10 mg/kg animal weight) hamster was introduced into the cloth cages for 1 hr to ensure adequate time for blood feeding, following the methods of Modi and Tesh (1983) for other phlebotomine species. After blood feeding, 100 females were separated into individual oviposition rearing chambers. Water and glucose (60%) were provided in cotton swabs placed on the muslin covering of each chamber. The rearing chambers were kept in two large Styrofoam boxes with a hygrometer and maximum-minimum thermometer. The rearing chambers measured 5.5 cm high and 6.6 cm in diameter with the bottom lined with 1.5 cm of plaster of Paris.
The larval diet for rearing was made according to Young et al. (1981) with the following changes: bovine manure and dog food instead of dry rabbit feces and Purina Rabbit Chow. It was stored for approximately three months and stirred every two weeks. RESULTS AND DISCUSSION DESCRIPTION OF THE DEVELOPMENT INSTARS Egg - The eggs of the Lu. shannoni were dark brown in color with polygonal sculpturing over the chorion similar to other species of Lutzomyia (Forattini 1973, Ferro et al. 1987). The eggs were elongated oval, approximately 310 um long and 95 um wide. The females required vertebrate blood for the maturation of the follicles, and the first oviposition began between four and eight days after blood ingestion. In the rearing chamber, a majority of the females oviposited on the humid layer of gypsum. However, some females oviposited on the glass walls of the rearing chamber; these eggs were removed to the gypsum surface with a fine brush. The average duration for the egg stage was 8.5 days with a range of 6 to 12 days (Figs 2, 3).
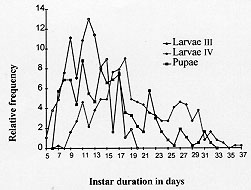
Figure 3: comparison of laboratory colony development times (in days) in the first three instars of Lutzomyia shannoni.
Feeding activity began within minutes after eclosion. The first stage larvae apparently preferred organic matter of plant origin, since they were observed devouring the small bits of grass in the manure. The first instar lasted an average of 9.6 days and a range between 5 and 13 days (Figs 2, 3). Second instar larvae - The color was similar to that of the first instar but this instar was larger in size, with four caudal setae approximately twice the body length. A Yform dorsal suture was present in the head capsule. The duration of this instar was from 4 to 13 days (ave. 9.2 days) (Figs 2, 3). Third instar larvae - The color was similar to that of the first two instars but it was larger, and again, it had four caudal setae and a Y-form dorsal suture in the head capsule. The duration of this stage ranged from 5 to 19 days (ave. 11.8) (Figs 2, 5).
Figure 5: comparison of third stage larvae, fourth stage larvae and pupae for number of days spent in each stage for colonized Lutzomyia shannoni.
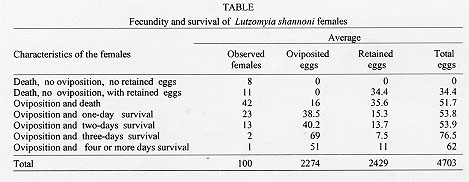
Pupa - Obtected, measured approximately 2600 um in length. It was glued to the substrate by the terminal abdominal segments consisting of the terminal pseudopodium of the ninth abdominal segment and the entire exuvium of the fourth instar larva. The pupal period ranged from 7 to 32 days (ave. 15.2 days) (Figs 2, 5). Adults - The longevity was between 4 and 15 days (ave. 8.6 days) ( Fig. 2). The total egg to adult period ranged from 36 to 74 days (ave. 54.6 days). Barretto, in a 1942 publication cited by Forattini (1973), presented the life cycle of Lu. shannoni at temperatures between 19.5 and 22.5 C as follows: egg stage ranging between 9 and 13 days, larva period ranging between 31 and 46 days, pupa period of 15 days and total range between 50 and 74 days. Comparing these results with the results found for Lu. shannoni in present study, we observed little variation in the egg stage, however, in the other stages of development we observed considerable variation. Fecundity analysis - As seen in the Table, 8% of the females neither oviposited nor retained eggs in spite of a full blood meal. Eleven percent of the females did not oviposit but retained an average 34.4 eggs. Females survival was proportional to numbers of eggs laid and inversely proportional to eggs retained. Since more than half of the eggs were retained by the females, the laboratory conditions probably were not sufficiently favorable for oviposition. Field-collected females of this species very rarely were found with retained eggs, and never more than one. On average, each female oviposited 22.7 eggs. The average egg retention per female was 24.3 eggs. The average total fecundity per female was of 47.0 eggs. This study is part of the research project "Genetics and Biogeography of Sand Fly Disease Vectors" in collaboration with the Instituto Nacional de Salud of Colombia and Yale University, USA, and funded by a U.S. National Institutes of Health Grant (AI-34521) to LEM and the Instituto Nacional de Salud of Colombia, project No. 3100302 of the Laboratory of Entomology. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98036d.jpg] [oc98036f.jpg] [oc98036e.jpg] [oc98036a.jpg] [oc98036c.jpg] [oc98036b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}