 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Metabolic Post-feeding Changes in Fat Body and Hemolymph of Dipetalogaster maximus (Hemiptera:Reduviidae) Lilian E Canavoso, Edilberto R Rubiolo*/^+
Departamento de Bioquimica Clinica, Facultad de Ciencias Quimicas, UNC,
Agencia Postal 4, CC 61, CP 5000, Cordoba, Argentina Received 21 July 1997; Accepted 15 January 1998
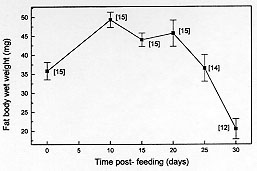
Code Number:OC98043 Lipids and glycogen in fat body as well as the modifications in the wet weight of this organ were evaluated in an unfed insect, Dipetalogaster maximus, on day 5 after adult ecdysis (time 0) and during a 30-day period after ingestion of blood meal. Total lipids, high density lipophorin (HDLp), carbohydrates, total proteins and uric acid were determined in the hemolymph during the same period. Fat body wet weight was maximum on day 10 post-feeding and represented on day 30 only 42% of the maximum weight. Lipids stored in the fat body increased up to day 15 reaching 24% of the total weight of tissue. Glycogen was maximum on day 20, representing approximately 3% of the fat body weight. HDLp represented at all times between 17-24% of the total proteins, whose levels ranged between 35 and 47 mg/ml. Uric acid showed at 20, 25 and 30 days similar levels and significantly higher than the ones shown at days 10 and 15. Hemolymphatic lipids fluctuated during starvation between 3-4.4 mg/ml and carbohydrates showed a maximum on day 15 after a blood meal, decreasing up to 0.26 mg/ml on day 25. The above results suggest that during physiological events such as starvation, the availability of nutrients is affected, involving principally the fat body reserves. Key words: carbohydrates - Dipetalogaster maximus - fat body - hemolymph - lipids - lipophorin - proteins The study of hemolymph components as well as of fat body deposits involved in essential metabolic processes related to supply of energy in insects for activities such as flight, reproduction, embryogenesis and metamorphosis provide important information about physiology of members of the same or different species (Wigglesworth 1972, Wyatt Pan 1978). Most animal species are able to overcome fasting periods and each species fits its metabolism into the particular situation of living, mobilizing its reserves when food is not available. However, among insects belonging to the same family, there is a remarkable difference in the possibilities of getting a food source resulting in distinct metabolic adaptation against fasting situations (Ziegler 1991). Several factors such as diet, temperature and disease influence the insect hemolymph composition of a species (Mullins 1985). The fat body, a tissue with multiple metabolic functions, performs important biosynthetic activity during the whole life of the insect and serves for nutrient storage (Keeley 1985). In reduviid bugs, hemimetabolous and obligately hematophagous insects, feeding and deposits in the fat body of metabolic reserves are important elements to cover their needs in the different stages as well as for their survival as adults (Friend Smith 1985). Nowadays, the information in fasted reduviids about biochemical changes in fat body as well as metabolic compounds related to supply of energy is rather scarce. This is an important aspect since the low weight, low weight/ length ratio of the insect (Schofield 1980, Lehane Schofield 1980, Schofield et al. 1992), a poor nutritional status (starvation) (Sjogren Ryckman 1966) and the need of a previous fasting period (Gringorten Friend 1979, Ward et al. 1982) have been pointed out in different species of reduviid bugs as factors stimulating flight. More recently, we demonstrated in vitro that three species of triatomine bugs have the biochemical constituents for generating partial transformation of high density lipophorin (HDLp) into low density ones (LDLp), employing fat bodies with more than 20 days of starvation (Canavoso Rubiolo 1994, 1995). This suggests the importance of the metabolic reserve levels available in the fat body during flight. This activity is an important mechanism of active dispersion by the vectors of Trypanosoma cruzi, responsible for American trypanosomiasis (WHO 1991). In the present study, we report the post-feeding modifications on fat body and hemolymph in a member of the Hemiptera order, Reduviidae family, the Dipetalogaster maximus. Adult insects fed 5 days after final ecdysis were submitted to different periods of starvation for 30 days. In fat body were analyzed the variations of glycogen, total lipids and the organ wet weight and in the hemolymph, the total lipids, HDLp, total proteins, uric acid and carbohydrates. MATERIALS AND METHODS Chemicals - All chemical reagents were of analytical grade. Doubly redistilled water was used throughout the experiments. Insects - Colonies of triatomine bugs D. maximus were reared and kept in the insectary, fed fortnightly on hens' blood. For the experiences, male grown-up insects were fed at day 5 after final ecdysis and separated in groups of 10-15 insects with similar weights. These groups were starved for 10, 15, 20, 25 and 30 days post-feeding and maintained at 28 C +/- 1 C , 60-70% humidity and 8:16 hr light:dark photoperiod as previously reported (Canavoso Rubiolo 1993). Groups of 10-15 insects in similar conditions and without meal were used at day 5 after final ecdysis in order to determine lipids and glycogen in fat body and the wet weight of the organ (time 0). Hemolymph collection - After 10, 15, 20, 25 and 30 days post-feeding, the hemolymph from the groups of insects was collected by centrifugation in the presence of 10 mM Na2EDTA and mixture of protease inhibitors (Canavoso Rubiolo 1993). The hemolymph samples were centrifuged at 10,000 g for 10 min at 4 C to remove hemocytes and stored at -70 C until assays were performed. For the carbohydrates determination, the hemo-lymph was collected only with Na2EDTA and immediately used. Chemical determinations in hemolymph - Protein concentrations have been determined by Bradford (1976), using albumin bovine as standard whereas uric acid was determined by enzymatic methods, employing a commercial kit (Uricostat, Wiener Lab., Argentina). Total lipid determinations were performed according to Frings and Dunn (1970) and carbohydrates, according to Scott and Melvin (1953). Determination of lipophorin HDLp Lipophorin standard - A standard curve for measurement of HDLp by ELISA was carried out employing HDLp from D. maximus, isolated from hemolymph by KBr density gradient ultracentrifugation and dialysed overnight against phosphate buffered saline (PBS) at pH 7.4 (Canavoso Rubiolo 1995). The concentration of lipophorin standard was quantified by Bradford (1976). ELISA assays - An indirect ELISA was performed in order to determine lipophorin titer (Voller et al. 1976). Microtiter plates (Labsystem) were loaded with 200 ml/well of the lipophorin standard (10-16 ng) or hemolymph from different times post-feeding diluted (1:300,000 to 1:1.000,000) with buffer (15 mM sodium carbonate, 35 mM sodium bicarbonate, pH 9.6) and incubated for 120 min at 37 C. Then, the microplates were washed five times with phosphate-buffered saline with Tween 20 (PBS-Tween) (8.2 mM sodium phosphate, 1.5 mM potassium phosphate, 150 mM sodium chloride, 2.7 mM potassium chloride, 0.05% Tween 20, pH 7.4). Serum anti-lipophorin HDLp from adults of D. maximus, obtained in rabbits as described previously (Canavoso Rubiolo 1995), was diluted in PBS-Tween (1:10.000) containing 0.1% bovine serum albumin and loaded into each well incubating for 60 min at 37^ C. Plates were washed and loaded with anti-rabbit inmunoglobulin conjugated to horseradish peroxidase (Sigma) in PBS-Tween and incubated for 30 min at 37^ C. Then, the plates were washed again and developed at 37^ C for 30 min with o-phenylenediamine in substrate buffer (81.5 mM sodium phosphate, 33.3 mM citric acid; 0.08% hydrogen peroxide, pH 5.0) and read at 490 nm. Determination of fat body wet weight - At time 0 (unfed insects, 5 days after final ecdysis) and after 10, 15, 20, 25 and 30 days post-feeding, fat bodies were carefully dissected, washed, dried on filter paper and weighed with a +/- 0.01 mg precision. Determination of lipids and glycogen in fat body - The lipids extraction was carried out according to Folch et al. (1957) and the material was taken up to constant weight in a N2 atmosphere. For determination of glycogen, the fat bodies were homogenized in a KOH 30%, boiled for 30 min and precipitated with ethanol (Hassid Abraham 1957). Then, the carbohydrates were determined by the anthrone method (Scott Melvin 1953). Statistical analysis - Student's t tests for observations were utilized in all instances. Results are reported as a mean +/- SEM. RESULTS Fat body wet weight of D. maximus after a blood meal changed with starvation compared to unfed insects (time 0), reaching 49.32 +/- 2.07 mg on day 10 and decreasing to 20.52 +/- 2.68 mg in insects fasting for 30 days (P < 0.01 day 30 vs day 10). The last value represented only 42% of the maximum wet weight reached. In the insects fasting during 25 days, the fat body wet weight was similar to that found in unfed animals (Fig. 1).
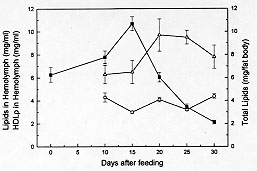
Figure 2: variations of lipids (-m-) and HDLp (-r-) in hemolymph and deposits of total lipids in fat body (-n-) at different times post-feeding. Values are mean +/- SEM (n= 10), determined in duplicate.
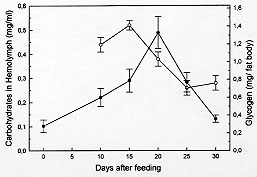
Previous to feeding of the insects, the glycogen reserves in fat body represented only 0.78% of the tissue (0.28 mg per fat body). This value is 5 and 1.3 folds lower than the maximum (day 20) and the value at day 30 post-feeding respectively (Fig. 3) whereas the concentration of carbohydrates in the hemolymph underwent important changes, dropping from 0.52 mg/ml to 0.26 and 0.28 mg/ml during the two last periods analyzed.
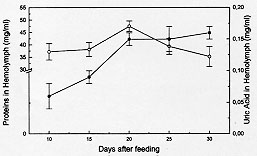
Figure 4: concentration of total proteins (-m-) and uric acid (-l-) in hemolymph of Dipetalogaster maximus at different times post-feeding. Values are mean +/- SEM (n= 10), determined in duplicate.
DISCUSSION Fat body in insects may be considered functionally, analogous to both vertebrate liver and adipose tissue, serving not only for the storage of reserves, but also as a general center for intermediary metabolism (Beenakkers et al. 1985). The hemolymph is the medium through which nutrients are transported from the sites of absorption directly to the sites of tissue respiration or to storage organs and, subsequently, to sites of utilization. Also, it transports waste products to excretory organs and all chemical exchanges between the organs affected (Wigglesworth 1972, Beenakkers et al. 1985). The present work reports about biochemical modifications occurring in the fat body reserves and in the hemolymph after a blood meal in the hemipteran D. maximus. They were evaluated in fat body through glycogen, total lipids and wet weight of the organ as well as by hemolymphatic levels of carbohydrates, total lipids, HDLp, proteins and uric acid. This hemimetabolous insect, a unique species of their genera, lives among rocks and feeds principally on blood lizards and wild mammalians (Ryckman Ryckman 1967, Beltran Carcavallo 1985). Despite the blood meal results critic for reduviids metamorphosis and that the nutritional reserve after moulting has an important effect on the development and growth (Friend Smith 1985), their ability to survive long periods without feed is well known (Szumlewicz 1976). Blood ingested by triatomine bugs is primarily stored in the anterior midgut where water absorption occurs quickly whereas the major digestion and absorption processes are relegated to posterior midgut (Terra Ferreira 1994). These events are influenced by amount of consumed blood, physiological status, sex and temperature (Catala et al. 1992). Our results indicated that after feeding D. maximus performs its digestion process accumulating reserves continuously up to day 20 post-feeding. In this period, the maximum value of the fat body wet weight was reached at day 10. At days 25 and 30 post-feeding, fat body wet weight showed an appreciable decrease, indicating that the mobilization process of reserves prevails over storage process, representing at day 30 less than 50% of the maximum weight (Fig. 1). However, the ratio between total weight of insect and fat body wet weight did not show significant changes through the times analyzed; the fat body being about 5% of the total weight of insect (data not shown). Deposits of total lipids and glycogen showed similar patterns, differing at times at which they reached the maximum values (Figs 2, 3). An interesting aspect was observed in glycogen content found at day 30, glycogen at this time being higher than the ones found in unfed insects (time 0). This phenomenon associated to lipidic deposits at time 0 and to nutritional reserves after ecdysis as pointed by Friend and Smith (1985) could suggest that these hematophagues employ the carbohydrates preferably at lipidic reserves, generating fuel to moulting as adult and reserving lipids for eventual flights in order to look for food. This speculation is supported by the higher energetic value of lipids compared to carbohydrates and the fact that lipids are generally the major fuel for flight used for insects (Beenakkers et al. 1985). Unlike glycogen, the lipidic deposits decreased markedly after day 15 post-feeding up to day 25, when they represented only 3.4 mg per fat body (Fig. 2). They showed triacylglycerols as a predominating lipid amongst non-polar components and phosphatidylcholine and phosphatidylethanolamine as the main polar components (Canavoso et al. 1996). Influence of starvation upon fat body weight and carbohydrates metabolism has been reported in the phytophagous hemimetabolous Carausius morosus. It was observed that glycogen content decreased after 15 hr of starvation and virtually disappears after 24 hr whereas fat body weight decreased during a 4-day period of starvation by about 30% (Lohr Gade 1983). Influence of starvation has also been analyzed in the holometabolous Manduca sexta by Ziegler (1991). Thus, it was observed after 5 days of starving a decrease in both, the fat body dry weight and in the lipidic content while glycogen reserves were exhausted. A similar pattern was demonstrated in adult locusts (Jutsum et al. 1975). It is possible therefore that the striking differences in carbohydrate deposits before described may be attributed to different processes of digestion between these hematophagous insects and phytophagues. On the other hand, levels of hemolymphatic carbohydrates extremely elevated or absent have been reported in insects and some arthropods (Bedford 1977). In this respect, our results obtained for D. maximus were lower compared to other members of the Insecta class: Periplaneta americana (Steele 1961), Locusta migratoria (Goldsworthy 1969), Acheta domesticus (Nowosielski Patton 1964) and C. morosus (Lohr Gade 1983). Since carbohydrates are used as fuel by insects in the initial flight period (Beenakkers et al. 1985), we believe that hemolymphatic carbohydrates may also be required by reduviids during the first phase of this activity. Preliminary experiments carried out in our laboratory with triatomine bugs submitted to flight, showed that a few minutes later the hemolymphatic carbohydrates were undetectable (data not shown). The insects mobilize the lipids from fat body as diacylglycerols (Beenakkers et al. 1985, Van der Horst et al. 1993). In D. maximus, the hemolym-phatic lipids showed a permanent fluctuation , possibly due to their energetic contribution to the vital functions of tissues (Fig. 2). Ward et al. (1982) have suggested that lipids in flight muscle increase probably at the expense of the fat body lipids. A similar pattern of changes between HDLp levels and hemolymphatic proteins was observed during post-feeding time (Figs 2, 4), reaching both the maximum values at day 20 post-feeding. Moreover, HDLp represented all time evaluated 17-24% of the hemolymphatic proteins. Our result differs from those found by Gonzalez et al. (1991) in Triatoma infestans in both, the levels HDLp and the proteins probably due to a different experimental design and the analytical procedure applied for the quantification of HDLp. On the other hand, it has been pointed out that in starved insects, the hemolymphatic proteins play an important role as reserve (Wyatt 1961). D. maximus showed increasing levels of circulating proteins with fasting up to day 20, probably as a consequence of the predominance of the mechanisms of synthesis and transport of absorbed proteins up to deposit organs upon their catabolism (Fig. 4). Uric acid is the principal nitrogenous excretory product of insects (Wyatt 1961), and more than 90% of the nitrogen excreted by fed Rhodnius prolixus is represented by this metabolite (Wigglesworth 1931). The levels of uric acid in hemolymph of D. maximus increased from day 10 (0.06 mg/ml) up to 0.16 mg/ml on day 30 (Fig. 4), in agreement with those reported by Barret and Friend (1966) employing a non-enzymatic method and starving fifth instar of R. prolixus. Since hemolymph volumes decline with time (Barret Friend 1966) it is controversial if the uric acid increase observed in D. maximus is due to an increase of proteic catabolism or results merely from a decrease of the hemolymph volume. In case this last assumption were correct, either uric acid would not be produced during starvation or if the uric acid is formed, the amount would be equal to their clearance. Since Wigglesworth (1972) has shown that stored proteins in fat body of R. prolixus disappear during starvation, the former suggestion appears more probable. During the period analyzed, hemolymphatic urea was not detected in D. maximus. For production of urea, arginase has been suggested to be involved in many species of insects (Cochran 1985). Apparently, D. maximus would lack arginase although Friend and Smith (1985) have reported the presence of urea in R. prolixus urine 2-3 hr post-feed, pointing to their probable exogen origin. We conclude that physiological events such as fasting affect the main metabolic processes in these insects. Future research, directed towards the qualitative and quantitative changes of the lipidic components from fat body induced by starvation, may lead to a better physiological and biochemical knowledge of these obligately hematophagous insects and could contribute to understanding the need of one poor nutritional status to begin the flight. In triatomine bugs, flight can take place when food sources are absent or inaccesible or when vectors colonize other geographic areas. Moreover, flight has been pointed out as responsible for the active dispersion of these insects, with implication on their control and on the American trypanosomiasis epidemiology. This work was supported by grants from SECyT- Universidad Nacional de Cordoba and CONICOR. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98043c.jpg] [oc98043a.jpg] [oc98043d.jpg] [oc98043b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}