 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Properties of Beta-lactamase from Neisseria gonorrhoeae Marta C de Castillo^+, Fernando Sesma, Olga M de Nader, Aida P de Ruiz Holgado
Instituto de Microbiologia, Facultad de Bioquimica, Quimica y Farmacia,
Universidad Nacional de Tucuman, Ayacucho 471, (4000) S.M. de Tucuman,
Argentina Received 11 August 1997; Accepted 15 January 1998
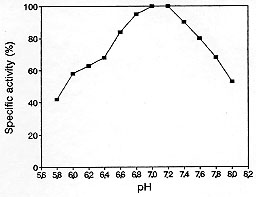
Code Number:OC98045 Beta-lactamase activity was studied in Neisseria gonorrhoeae strains. Optimum temperature was found to be 37 C. The enzyme was inactivated at temperatures higher than 60 C, but remained active during storage at low temperatures (4 C, -30 C and -70 C) for two months. Enzyme activity was observed within a pH range of 5.8-8.0, while the optimum pH was 7.0-7.2. Addition of Ni^2+, Fe^2+, Fe^3+, Mn^2+ and p-chloromercurybenzoate to the reaction buffer exerted a negative effect upon the activity, whereas Hg^2+ and ethylene diamine tetra-acetic acid produced complete inhibition. These results would indicate the presence of -SH groups at the catalytic site of the enzyme. Key words: Neisseria gonorrhoeae - beta-lactamase - enzyme activity Penicillin became the drug of choice for treatment of infections caused by Neisseria gonorrhoeae shortly after it became available for clinical use. Remarkably small doses (e.g., 100,000 units altogether) were used to cure gonorrhoea (Mahoney et al. 1945), an occurrence reflecting the high susceptibility of the gonococcus to penicillin (Reyn et al. 1958). During the subsequent decades a gradual increase in gonococcal resistance to penicillin, documented in the United States (Martin et al. 1970) and elsewhere (Amies 1969, Reyn 1961, 1969, Arya & Phillips 1979), needed increases in the dose of penicillin as well as the addition of probenecid to maintain effective therapy (Holmes et al. 1973, Kaufman et al. 1976). In the United States, however, a reduction in gonococcal resistance to penicillin has been witnessed since 1972 (Reynolds et al. 1976), and in 1976 a substantial decline in the rate of increase in morbidity from gonorrhoea was observed (CDC 1977). The isolation of penicillinase-producing N. gonorrhoeae (PPNG) (Ashford et al. 1976, Phillips 1976) urged the use of other antimicrobials. The first two reported cases of infection with PPNG in the United States were in March 1976 in Maryland and in April 1976 at Travis Air Force Base in California (CDC 1976). In Argentina the isolations of PPNG strains reached a value of 30% in 1995 (Mernes et al. 1997). The present work was carried out in order to characterize and to study the properties of beta-lactamase from N. gonorrhoeae as to contribute useful information. MATERIALS AND METHODS Microorganisms - Two strains of N. gonorrhoeae, isolated from clinical specimens, were used. The strains were maintained at -70 C in a cryoprotective medium (Tryptic Soy Broth -BBL- with 25% glycerol). Strains were selected according to their capacity to produce beta-lactamase. Preparation of cell-free extracts - Portions of SJ-GC broth (not commercially avilable) (500 ml) were inoculated and incubated at 37 C for 24 hr (Shockley et al. 1980). Subsequent operations were carried out between 0 and 4 C. Cells were harvested at the end of the log phase by centrifuging at 6,000 g for 20 min, washed twice with phosphate buffer (pH 7.0) and resuspended in the same buffer to give a final concentration of 30% wet weight (w/v). Cell suspensions were disintegrated in a French press at a constant pressure of 1,575 kg cm^-2. Cell debris was removed by centrifuging at 30,000 g for 10 min. The supernatant fluid was used as crude enzyme extract. Enzyme assays - beta-lactamase activity was determined by a direct spectrophotometric method (O'Callaghan et al. 1972). Haemophylus influenzae ATCC 10211 was used as a negative control, whereas Escherichia coli ATCC 35218 was used as a positive control. Hydrolysis of the beta-lactamic ring was measured by the decrease of optic density of the cephaloridine solution. Measurements were carried out at 255 nm. One enzyme unit (U) was defined as the amount of enzyme that released 1 umol of cephaloridine per ml per min under the given assay conditions. Specific activity was expressed as enzyme units per mg of protein. Protein concentration was determined by the method of Lowry et al. (1951) with bovine serum albumin (BSA) as a standard. Isoelectric focusing - Electric focusing was performed on a sucrose ampholyte gradient (pH 3.0 to 10.0). The experiment was carried out below 4 C on an ampholyte electric focusing column (LKB 8100) for 48 hr with a final gradient of 300 U. The contents of the column were cut out into 3 ml fractions and each fraction was assayed for its beta-lactamase and pH (Matthew et al. 1975). Determination of optimal pH, temperature and thermal stability - The effect of the pH on the beta-lactamase activity was determined in phosphate buffer (0.2 mol l^-1) with a pH range of 5.8-8.0. The influence of the temperature on the enzymatic activity was determined by incubating the assay mixture for 15 min at temperatures ranging from 15 C to 55 C. Thermal stability was determined by incubating the enzyme extract at temperatures ranging from 15 C to 60 C for 1 to 25 min. Substrate was then added and the solution incubated for an additional 15 min at 37 C, to measure the residual activity. Storage stability of the enzyme - Enzyme suspensions were stored at 25 C, 4 C, -30 C and -70 C for 1, 2, 5, 10, 20, 30, 45 and 60 days. The residual enzyme activity was determined after each period. Effect of metal ions, inhibitors and other substances on enzyme activity - Stock solutions of CaCl2, HgCl2, CuSO4, MnCl2, FeCl3, CdCl2, NiSO4, ZnSO4, MgCl2 and FeSO4 were prepared in phosphate buffer (pH 7.0), and added separately to the reaction mixture in order to obtain a final concentration of 10^-2, 10^-3, 10^-4 and 10^-5 mol l^-1. All reagents were purchased from Sigma. Inhibitors such as p-chloromercurybenzoate (PCMB) (Sigma) and ethylene diamine tetra-acetic (EDTA) (Sigma) were assayed at the same concentrations as the metal ions. Residual enzyme activity was assayed and expressed as a percentage of the activity determined in phosphate buffer (control). Reproducibility - All results presented in this paper are the means of three replicate assays. RESULTS AND DISCUSSION The two beta-lactamases isolated from two strains of N. gonorrhoeae in this study showed similar behaviour, indicating that they were similar. Therefore further studies were carried out with only one of the isolates. Under the assay conditions used in this study cell-free extracts of N. gonorrhoeae showed an activity of 0.36 U per mg of protein. The isoelectric point obtained was 5.4, which suggests that this beta-lactamase isolated from N. gonorrhoeae belongs to the TEM-1 type (Van Embden et al. 1980). Percival et al. (1977) and Phillips (1976) found similar results in other Neisseria strains. The effect of the pH on the enzyme activity is shown in Fig. 1. Optimum pH was found to be 7.0-7.2. These results agree with those reported by Livermore and Corkill (1992) who found that pH values 6.0-7.0 were optimal for beta-lactamase activity in Escherichia coli 976 (beta-lactamase TEM-1).
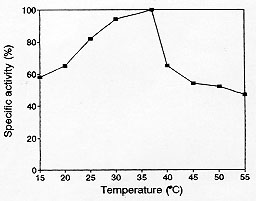
Figure 2: effect of the temperature on the specific activity (U/mg of enzyme) of beta-lactamase from Neisseria gonorrhoeae. Temperatures assayed: 15, 20, 25, 30, 37, 40, 45, 50 and 55 C.
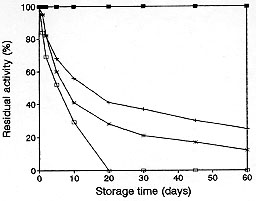
The enzyme solutions showed a remarkable stability when kept at low temperatures. The residual enzyme activity of the extracts remained constant for two months throughout storage when kept at -70 C ( Fig. 3). The enzyme was totally inactivated at 25 C after 20 days of storage. Similar results have been obtained by Edwards and Greenwood (1990) for beta-lactamase isolated from Bacteroides, in which case the enzyme retained 100% activity after storage at -70 C for 10 months. The stability of beta-lactamase at low temperatures as found in this work will prove to be of great importance when enzyme purification studies are undertaken.
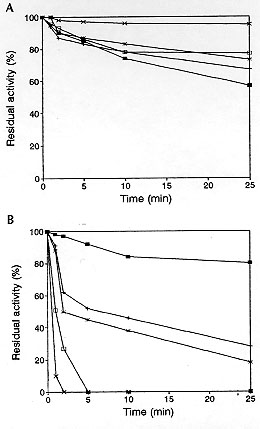
Figure 4: thermal denaturalization of beta-lactamase activity from Neisseria gonorrhoeae. Enzyme extracts were incubated at the temperatures mentioned below for 1, 2, 5, 10 and 25 min. Residual activity was measured after substrate was added and the solution incubated for an additional 15 min at 37 C. A: 15 C (n), 20 C (+), 25 C (*), 30 C (o) and 37 C (x). B: 40 C (n), 45 C (+), 50 C (*), 55 C (o) and 60 C (x).
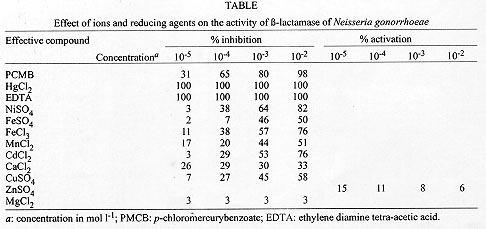
The Table shows the effect of cations on the enzyme activity. Among the cations examined, Hg^2+ showed complete inhibition of the enzyme at all assayed concentrations, whereas Ni^2+ caused an 82% decrease of the enzyme activity at the highest studied concentration (10^-2 mol l^-1). Hg^2+ is a strong inhibitor of several beta-lactamase enzymes isolated from different sources, a fact that suggest the presence of thiol groups at the catalytic site. Similar results have been reported by Hirai et al. (1980) who worked with Pseudomonas cepacia and found a 100% loss of the enzyme activity at a concentration of 3x10^-3 mol l^-1 of the cation, and by Fujii et al. (1986) for Legionella gormanii. The addition of other divalent cations, e.g. Fe^2+ and Ni^2+, to the reaction buffer affected the enzyme activity too. The presence of chelating agents such as EDTA inhibited beta-lactamase activity 100%, suggesting that a metal cation is required for its activity. Similar results have been observed by Saino et al. (1982) working with Pseudomonas maltophilia. Addition of PCMB to the reaction mixture produced a marked inhibition (about 98%) of beta-lactamase activity from N. gonorrhoeae. The loss of enzyme activity in the presence of PCMB, an agent that oxidizes -SH groups, also supports the idea of the involvement of -SH groups in the beta-lactamase activity. The inhibitory effect exerted by PCMB has been reported in earlier studies for strains of P. cepacia (Hirai et al. 1980), Bacillus (Sabath & Finland 1968), Proteus (Imsande et al. 1970) and for Bacteroides fragilis (Yotsuji et al. 1983). Addition of Zn^2+ at a concentration of 10^-5 mol l^-1 produced a slight stimulation of the enzyme activity, 15%, which agrees with the results found by Sabath and Abraham (1965) for B. cereus. The results obtained in the present work allowed the identification of beta-lactamase in N. gonorrhoeae. As the properties of beta-lactamase of different bacteria vary extremely it is important to study them in each strain. Specific enzymes which inhibit penicillin have been studied and classified. Their properties have been studied in crude extracts from Proteus, Klebsiella , Enterobacter, E. coli, Pseudomonas (Alyffe 1965) and Bacteroides (Edwards & Greenwood 1990). Further investigations are necessary to obtain a complete characterization of the beta-lactamases and to be able to classify them, determining their kinetic properties and behaviour with respect to physical and chemical agents. This work provides information that will give us optimal treatments as a benefit, which are important for the implementation of elemental sanitary policies. ACKNOWLEDGEMENTS To Mr Eric Fengler for language advice and drawing of the graphics. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98045b.jpg] [oc98045a.jpg] [oc98045d.jpg] [oc98045c.jpg] [oc98045e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}