
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 93, Num. 2, 1998, pp. 243-246
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 93 (2), March 1998, pp. 243-246
RESEARCH NOTE
Structure of the Knob Protein Gene of the Saimiri Monkey-adapted
Palo Alto Strain of Plasmodium falciparum
Wanda Chalvet, Jean-Francois Pouliquen*, Thierry Fandeur/^+
Laboratoire de Parasitologie Moleculaire
*Laboratoire des Retrovirus, Institut Pasteur de la Guyane, BP 6010, 97306
Cayenne Cedex, French Guiana
^+Corresponding author. Fax: +5-94-31.8083
Received 20 September 1997; Accepted 8 January 1998
Code Number:OC98046
Sizes of Files:
Text: 11.3K
Graphics: Line drawings and photographs
(jpg) - 145.8K
Key words: Plasmodium falciparum - histidine-rich protein - Palo
Alto parasites - squirrel monkey - strain infectivity
RESEARCH NOTE
The histidine-rich protein is the major component of the knob-like
protrusions detected on the membrane of both human and monkey erythrocytes
infected with Plasmodium falciparum (SA Luse & LH Miller 1971 Am
J Trop Med Hyg 20: 655-660, M Hommel et al. 1982 Parasite Immunol
4: 409-419, F Ardeshir et al. 1987 EMBO 6: 1421-1427). The knobs
on the red blood cell surface are generally associated with strain
pathogenicity and virulence. Isolates collected from naturally infected
patients are consistently of knobby phenotype (Hommel et al. loc.
cit.) and knobless lines were reported to be less virulent in monkey
models (S Langreth & E Peterson 1985 Inf Immun 47: 760-766, T
Fandeur et al. 1992 Inf Immun 60: 1390-1396).
Experimental infection of the squirrel monkey, Saimiri sciureus,
with P. falciparum is one of these well-established monkey models
for human malaria. Recently, we examined several monkey-adapted and
culture-derived P. falciparum strains for their ability to propagate
in splenectomized Saimiri monkeys. Consistent with the above, we
found a correlation between allelic diversity at the HRPI locus and the
course of blood stage infection (T Fandeur et al. 1996 Exp Parasitol
84: 1-15). A particular allele of the HRPI gene was found only in
strains presenting a FUPSP genotype and growing to high parasitic density.
However, strains possessing a wild-type HRPI gene (FUPCB, FVO, Geneva, Sal
I, Honduras and FCH4) or lacking the HRPI sequences (FUPCP), produced no to
low-density infections in splenectomized Saimiri, thereby suggesting
that this particular HRPI allele is fundamental for the P.
falciparum infection of S. sciureus erythrocytes. We now report
the partial sequence of the HRPI allele which was associated with rapid
developing infection in the splenectomized Saimiri.
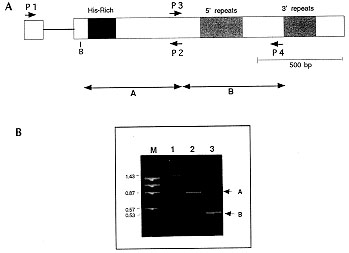
Figure 1: A - Structure of the HRPI gene of
Plasmodium falciparum. Boxed areas indicate the coding regions. 5'
and 3' repeats, and the histidine-rich regions are shaded. The positions of
the oligonucleotides P1-P4 used for the PCR reactions, and the positions of
PCR-fragments A and B sequenced after transfer to M13 are indicated. The
BamHI (B) restriction site near the N-terminal histidine-rich domain of
the gene is shown. Oligonucleotides P3 and P4 are identical to those
previously labeled HRPI A and D in an other study (T Fandeur et al. 1996
Exp Parasitol 84: 1-15), B - PCR-products generated with
oligonucleotides P1-P2 (lane 1) and P3-P4 (fragment B, lane 3) by using
FUPSP DNA as template. The 1.4 kb PCR product amplified by using primers P1
and P2, and digested with BamHI-EcoRI (lane 2).
The HRPI gene shown in Fig. 1a includes a region encoding a histidine-rich
portion in the second exon, and two blocks of repeated sequences. The gene
in FUPSP genomic DNA was amplified with primers P1 (CCGGGATCCATGAAAAGTTT
TAAGAACAA, positions 629 to 657) and P2 (TGAATTCCCTGCACCATGGGGTGGG,
positions 1597 to 1621), and with primers P3 (CCGGATCCCACCCCATGGTGCAGGC,
positions 1590 to 1614) and P4 (AGAATTCCATT GTCCTTTATTTGTTGCGGC, positions
2216 to 2245), as deduced from the HRPI sequence in FCR3 parasites (LG
Pologe et al. 1987 Proc Nat Acad Sci, USA 84: 7139-7143). The
artificial BamHI or EcorI sites introduced at the 5' end of
the primers are underlined. Priming the HRPI sequences with
oligonucleotides P1-P2 amplified a 1430 bp PCR-product (Fig. 1B, lane 1),
corresponding to a wild-type HRPI sequence, whereas the 530 bp fragment
produced by using primers P3-P4 (Fig. 1B, lane 3) is shorter than expected
from the previously published FCR3 and NF7 sequences (Pologe et al. loc.
cit., T Triglia et al. 1987 EMBO 6: 1413-1419). Indeed,
amplification of an intact 640 bp fragment was uniformly observed in FUPCB,
FVO, Geneva, Sal I, Honduras and FCH4 parasites, all producing low-density
and self-cured infections in splenectomized monkeys (Fandeur et al. loc.
cit.). The fragments labeled A and B in Fig. 1 were further
characterized by cloning and sequencing. The PCR-products were restricted
with BamHI-EcorI, excised from the gel, purified by Geneclean
II^R, and finally cloned into BamHI-EcorI digested M13
vectors. Digestion of the 1430 bp fragment (Fig. 1B, lane 1) produced two
restriction fragments of about 870 and 570 bp respectively ( Fig. 1B, lane
2). The BamHI restriction fragment located in the 5' region and
containing the intron was not further studied.
Figure 2: partial nucleotide sequence of the HRPI gene
from the FUPSP strain of Plasmodium falciparum adapted to the
Saimiri monkey, and comparison to those reported previously for the
FCR3 and NF7 strains. The sequences were aligned using the GeneWorks
program (Intelligenetics). The consensus sequence is shown together with
differences between the sequences; identities are shown as ( ..... ) and
deletions as ( ----- ). Unless specified otherwise, the positions of the
bases or amino acids given in the text are based on the numbering of this
consensus sequence. This sequence has been submitted to EMBL Nucleotide
Sequence Database with the accession number Y10828. The positions of PCR
fragments A and B shown in Fig. 1 are indicated.
The nucleotide sequences obtained of several independent bacteriophages
carrying HRPI fragments from FUPSP strain were determined (Fig. 2). The
sequences of the PCR-amplified regions A and B of the gene (Fig. 1A) were
joined end to end, and aligned to those previously published for the FCR3
and NF7 strains. The HRPI gene of FUPSP was closely related to that of FCR3
because both sequences lacked the 13 bases at positions 70-75, 102-107 and
1271 in the consensus sequence. Single base deletions at positions 848,
850, 859 and 1192 were detected only in the NF7 sequence. FUPSP and FCR3
sequences differed from each other by (i) seven point mutations
substituting Q, D, L, G, K, L, E in FCR-3 for H, N, P, D, E, S, Q in FUPSP
at positions 90, 99, 254, 755, 1267, 1287 and 1348, respectively; and by
(ii) a large deletion of 81 bp (1111 to 1192) in the FUPSP sequence.
Although the HRPI gene is generally well conserved in these isolates, the
few base substitutions and deletions have consequences on the encoded
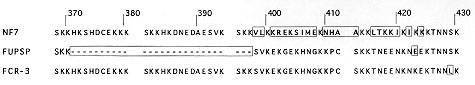
proteins. The deduced amino acid sequences differed mostly in the region
between from amino acid residues 370 and 440 (Fig. 3). A frame shift of NF7
occurs twice. The deletion of an A at position 1192 in the NF7 nucleotide
sequence modifies the reading frame from codon 398, it was then corrected
at position 424. No similar change in the reading frame was observed for
the FUPSP sequence despite a large deletion removing the fourth and fifth
5' repeats. Comparison of the predicted secondary alpha-helix and
beta-sheet structures of the protein in NF7, FUPSP and FCR3 indicated that
such changes result in altering the conformation of this region (data not
shown), in other respect described as highly immunogenic (MA Rashid et al.
1990 Mol Biochem Parasitol 38: 49-56), and also identified as being
the functional domain of the knob protein (A Kilejian et al. 1991 Mol
Biochem Parasitol 44: 175-182). These modifications are expected to
have some effect on the antigenic and functional properties of HRPI in the
context of FUPSP parasites.
Figure 3 alignment of predicted amino acid sequences
extending from residues 370 to 430 in strains NF7, FUPSP, and FCR3. Unique
residues, stretches of sequence, and deletions different between the
various alleles specificities of the strains are boxed. Amino acid
positions are deduced from the consensus nucleotide sequence shown in Fig.
2.
A general problem encountered when studying parasite virulence is to define
what virulence means. There is no single definition both satisfactory and
relevant for all situations studied. During infection of the splenectomized
monkeys with the various P. falciparum strains, we did not detect
any sign of severe disease, except for hyperparasitemia. In addition,
trophozoite and schizonte stages do circulate in splenectomized animals,
indicating that in this host low-grade parasitemias do not rely to
extensive parasite sequestration to capillary endothelium. Consequently, we
considered appropriate to assess strain virulence by monitoring
parasitemia. Based on this criterion, a correlation was established between
allelic diversity at the locus encoding the knob protein and the severity
of blood infections.
The positive selection in splenectomized monkeys for parasites expressing a
particular form of the knob protein is puzzling. By analogy to in
vitro observations (JG Culvenor et al. 1987 Exp Parasitol
63: 58-67), we speculate that adaptation to Saimiri, usually
associated with increased growth rate, might involve selection of a
parasitic strain with this particular allelic form of the HRPI gene.
However, we do not understand at present why there is positive selection
for these parasites. The function of HRPI remains unknown, yet some
evidence has been presented for the knob protein being a malarial parasite
analogue of red cell protein 4.1 (Kilejian et al. loc.cit.). This
suggests a mechanism whereby HRPI could modify the host cell membrane and
directly interfere with parasite development within the infected
erythrocyte. The diversity found at the HRPI locus might therefore reflect
spectrin diversity of the red blood cell as both proteins associate with
the erythrocyte membrane cytoskeleton to form knob complexes.
Acknowledgments: to Bernard Bonnemains for his assistance in
managing the monkeys and to Cecile Le Scanf for fruitful discussions.
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc98046a.jpg]
[oc98046b.jpg]
[oc98046c.jpg]
|

{kind=link}
{kind=link}
{kind=link}