 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Molecular and Antigenic Characterization of the Leishmania (Viannia) panamensis Kinetoplastid Membrane Protein-11
Jose R Ramirez, Christof Berberich/^+, Andres Jaramillo, Carlos Alonso*,
Programa de Estudio y Control de Enfermedades Tropicales, Sector de
Inmunologia y Biologia Molecular, Facultad de Medicina, Universidad de
Antioquia, AA 1226, Medellin, Colombia Received 29 October 1997; Accepted 15 January 1998
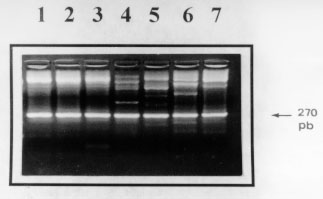
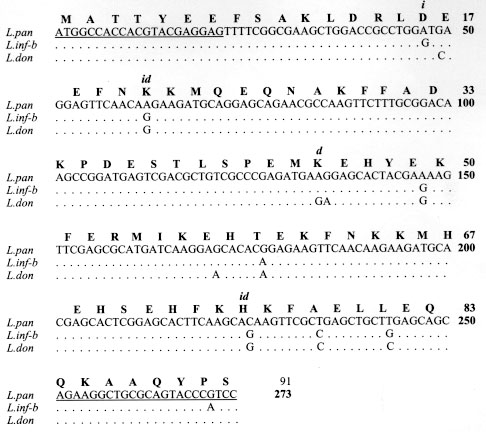
Code Number:OC98047 The kinetoplastid membrane protein 11 (KMP-11) has been recently described in Leishmania (Leishmania) donovani as a major component of the promastigote membrane. Two oligonucleotide primers were synthesized to PCR-amplify the entire coding region of New World Leishmania species. The Leishmania (Viannia) panamensis amplification product was cloned, sequenced and the putative amino acid sequence determined. A remarkably high degree of sequence homology was observed with the corresponding molecule of L. (L) donovani and L. (L) infantum (97% and 96%, respectively). Southern blot analysis showed that the KMP-11 locus is conformed by three copies of the gene. The L. (V) panamensis ORF was subsequently cloned in a high expression vector and the recombinant protein was induced and purified from Escherichia coli cultures. Immunoblot analysis showed that 80%, 77% and 100% sera from cutaneous, mucocutaneous and visceral leishmaniasis patients, respectively, recognized the recombinant KMP-11 protein. In a similar assay, 86% of asymptomatic Leishmania-infected individuals showed IgG antibodies against the rKMP-11. We propose that KMP-11 could be used as a serologic marker for infection and disease caused by Leishmania in America. Key words: New World leishmaniasis - Leishmania (Viannia) panamensis - kinetoplastid membrane protein-11 - antigenicity The number of cases of American tegumentary leishmaniasis (ATL) has been estimated to be around 59,000 every year, and approximately 59 million people reside in areas where the ATL-causing Leishmania species are transmitted (Ashford et al. 1992). ATL caused by the species complex Leishmania (Viannia) braziliensis is endemic in large areas of Central and South America (Grimaldi et al. 1989). In Colombia, L. (V) panamensis is responsible for most of the ATL cases, including cutaneous leishmaniasis (CL), mucocutaneous leishmaniasis (MCL) and disseminated cutaneous leishmaniasis (DCL) (Corredor et al. 1990, Colombian Health Ministry 1994, Velez et al. 1994). Control of leishmaniasis in the American continent is particularly complicated due to the fast evolving variety of Leishmania parasites and the complexity in their epidemiological patterns (Grimaldi & Tesh 1993). Recent attempts to develop molecularly defined vaccines for leishmaniasis have focused on major parasite cell surface molecules, including the metalloproteinase GP63, the promastigote surface antigen complex (PSA) and the lipophosphoglycans (LPG) (McConville et al. 1987, Chang et al. 1990, Burns et al. 1991). For reasons such as its abundance on promastigotes and its important physiological role in parasite-macrophage interactions (Turco & Descoteaux 1992), the latter was considered an attractive vaccine candidate. In early works, it has been demonstrated that immunization with LPG might confer immunoprotectivity in mice (Handman & Mitchell 1985, McConville et al. 1987, Russell & Alexander 1988) and that peripheral blood mononuclear cells from leishmaniasis patients were strongly stimulated to proliferate in response to LPG (McConville et al. 1987, Jaffe et al. 1990, Mendoca et al. 1991). However, it could be recently shown that T-cell stimulation associated with protective immunity elicited in mice, was due to a tightly associated small protein in the LPG fraction which was recently renamed as the kinetoplastid membrane protein-11 (KMP-11) (Jardim et al. 1991, Stebeck et al. 1995). So far, KMP-11 has been cloned and partially characterized in L.(L) donovani (Jardim et al. 1995a,b) and the L. (L) infantum and Trypanosoma brucei rhodesiense counterparts have been recently reported (Stebeck et al. 1996, Berberich et al. 1997). To date, no homologous locus in any New World Leishmania species has been isolated and characterized. Here, we report the cloning, sequencing and bacterial expression of the recombinant L.(V) panamensis KMP-11 protein. In addition, in a detailed study with sera from patients suffering from different forms of New World leishmaniasis we provide evidence that specific antibodies against KMP-11 are produced during natural course of infection. MATERIALS AND METHODS Parasites - Promastigotes of L. (V) panamensis (MHOM/PA/71/LS94), L.(V) panamensis (MHOM /CO/87/UA140), L. (V) guyanensis (MHOM/BR/75/M4147), L. (L) major (MHOM/MA/81/LEM265), L. (V) braziliensis (MHOM/CO/88/UA301), L. (V) colombiensis (IHAR/CO/85/CL500), and L. (L) infantum (MHOM/FR/78/LEM75), were grown at 26 C in Schneider`s Drosophila Medium (Sigma, USA) supplemented with 15% heat-inactivated fetal bovine serum and antibiotics (streptomycin 0.01 mg/ml, penicillin 10 U/ml). Promastigotes were grown to late logaritmic phase, harvested, washed three times with PBS and resuspended at 5 x 10^7/ml. Monoclonal and polyclonal antibodies - The L98 monoclonal antibody (mAb), originally raised against the L. (L) donovani LPG molecule (Tolson et al. 1989), but further described to recognize a carboxy-terminal epitope on the tightly associated KMP-11 molecule (Jardim et al. 1991, 1995a), was used to demonstrate the presence of the recombinant KMP-11. This mAb detects the 11 kDa protein in all kinetoplastid parasites so far tested (Stebeck et al. 1995). A rabbit polyclonal serum recognizing the recombinant L. (L) infantum KMP-11 (Berberich et al. 1997) was also used to detect the recombinant L (V) panamensis KMP-11 protein. Study subjects - Twenty six patients with parasitological diagnosis of leishmaniasis were studied. The presence of Leishmania parasites in patients was determined by direct microscopic examination and culture methods. Indirect immunofluorescence microscopy using mAbs capable to distinguish between species-specific LPG epitopes (McMahon-Pratt & David 1981) was applied in order to confirm the identity of Leishmania parasites. Twenty four patients had ATL caused by L. (V) panamensis or L. (V) braziliensis (15 CL and 9 MCL) and live in recognized endemic areas for leishmaniasis (departments of Antioquia, Tolima, Caldas and Cundinamarca, Colombia). Two patients had visceral leishmaniasis (VL) caused by L. (L) chagasi/L.(L) infantum and live in San Andres de Sotavento, an endemic region for VL at the Colombian Carribean Coast. ATL patients were treated as recommended by the WHO with Glucantime, 20 mg of antimony/kg daily intramuscularly (20 days for CL patients and 28 days for MCL patients) (WHO 1990). The two VL patients were treated with liposomal amphotericin B, 2 mg/kg intravenously for 21 days. A second group consisted of seven asymptomatic Leishmania-infected individuals that live in endemic areas for leishmaniasis and show a positive Montenegro skin test (MST). These individuals were clinically evaluated to detect any scar or current leishmaniasis compatible lesion. As negative controls, nine healthy individuals which have never lived or visited endemic areas for leishmaniasis and have a negative MST were included. Serum samples were collected from these individuals in sterile vacutainer blood collection tubes (Becton, Dickinson and Company, Rutherford, NJ, USA) and frozen at -20^ C until use. Polymerase chain reactions (PCR) - Genomic DNA was isolated as described previously (Medina-Acosta & Cross 1993). To amplify the entire coding region of the KMP-11 protein from the genomic DNA, the following oligonucleotide primers were used: forward, (CB1): 5" ATGGCCACCACGTACGAGGAG 3" and reverse, (CB2): 5" GGACGGGTACTGCGCAGCC TT 3". The primer sequences were based on the published KMP-11 coding region from L. (L) donovani (Jardim et al. 1995b). A "DNA thermal cycler 480" and "Amplitaq kit" (Perking-Elmer Cetus Instruments, New Jersey, USA) were used according to manufacturers instructions. Reactions mixtures included 25 pmol of each primer, 200 mM of each dNTP and 1.5 uM MgCl2. The following PCR protocol was performed: (a) denaturation at 94 C for 1 min; (b) annealing at 58 C for 1 min; and (c) extension at 72^ C for 1 min; 30 cycles. Southern blot analysis - 3 mg of L. (V) panamensis promastigote genomic DNA were digested with restriction enzymes (Sal I, Nco I, Xho I, EcoR I, Hind III and BamH I), separated on a 0.8% agarose gel, transferred to a nylon membrane (Hybond-N, Amersham Int., UK) and subsequently hybridized with an [alpha-^32P]dATP random-primed-labelled probe consisting of the 273 bp PCR fragment of the KMP-11 coding region. Hybridization was done in 6X SSC (1X SSC is 0.15 M NaCl, 15 mM Sodium-Citrate, pH 7.0), 0.1% SDS, 250 ug/ml hering sperm DNA and 50% formamide, at 42 C overnight. After hybridization, membranes were washed with (a) 2X SSC, 0.1% SDS; (b) 1X SSC, 0.05% SDS and (c) 0.1X SSC, 0.02% SDS, each washing step for 30 min at 42 C. Membranes were then exposed to an autoradiographic film for 96 hr. Cloning and sequencing - The 273 bp PCR fragment was blunt-end cloned in the EcoR V site of pBluescript (Stratagene, La Jolla, CA) and eight different recombinant clones were selected for restriction mapping to detect full-length inserts. Two of these clones were subsequently selected (clones pBx-KMP-4 and pBx-KMP-5) for DNA sequencing on double-stranded DNA by the dideoxy chain termination method (Sanger et al. 1977), using both comercial T7 and T3 primers or the CB1/CB2 primer pair and the "sequenase kit" (US Biochemical Corp., Cleveland, OH, USA). The analysis of DNA sequence was done using the University of Wisconsin Genetics Computer Group Program (Devereux et al. 1984). Subcloning, expression and purification of the recombinant protein - The pBx-KMP-5 insert was subcloned directionally into the BamH I/Hind III site of the expression plasmid pQE30 (Quiagen GMBH, Hilden, Germany). Induction of expression (2 mM IPTG) and protein purification was performed following the instructions of the manufacturer. Purification of proteins was performed under denaturing conditions in 8 M Urea and passage of the bacterial extract through a Ni-NTA affinity column. Purified protein was subsequently dialysed against 1X PBS. The expression and purification process of the recombinant protein was monitored by SDS-PAGE and coomassie blue staining. Western blotting - The purified recombinant protein was separated by 15% SDS-PAGE and electrotransferred to nitrocellulose membranes (Schleicher & Schöll Inc., Keene, NH, USA) using the MiniTrans blot system (BioRad Labs, Hercules, CA, USA). Protein transfer was monitored by Ponceau-S (Sigma Co., St Louis, USA) staining before blocking the filters in 5% blotto in PBS (Blocking solution) at 4 C overnight. The filters were then incubated with monoclonal (1:500), polyclonal (1:2000) or human (1:100) serum diluted in blocking solution with 0.5% tween-20 for 2 hr. To detect specific antibodies in human sera, the membranes were incubated successively with biotin-labelled anti-human IgG (Sigma Co., St Louis, USA) for 1 hr, streptavidin (BioRad Labs, Hercules, CA, USA) for 1 hr, and alkaline phosphatase-labelled biotin (BioRad Labs, Hercules, USA) for 1 hr. The immune reactions were visualized by incubating the membranes in bromo-chloroindolylphosphate-nitro blue tetrazolium (BCIP-NBT) for 30 min. For monoclonal and polyclonal sera, a peroxidase-labelled anti-mouse IgG and a alkaline phosphatase-labelled anti-rabbit IgG were used. Colour development was done with diaminobenzidine (DAB) and BCIP-NBT respectively. RESULTS Locus characterization and sequence of open reading frame - By using the CB1/CB2 primer pair, which sequence was based on the L. (L) donovani KMP-11 open reading frame (Jardim et al. 1995b), we amplified a 273-bp fragment in all species tested (Fig. 1) including L. (V) panamensis, L. (V) braziliensis and L. (V) guyanensis, the most important species causing CL and MCL in Colombia (Corredor et al. 1990, Colombian Health Ministry 1994). These results confirm, at the genomic level, the high degree of conservation for this gene between Old and New World Leishmania species as it was expected in view of the immunological cross-reactivity of the protein in various kinetoplastid species when using the L98 and L157 mAbs originally raised against the LPG (Tolson et al. 1994).
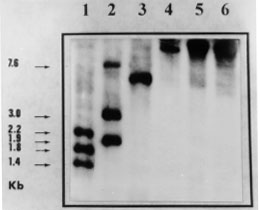
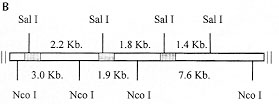
A: southern blot analysis of restriction enzyme digested genomic DNA using as probe a 273 bp-fragment of the KMP-11 coding region. Restriction enzymes: 1: Sal I, 2: Nco I, 3: Xho I, 4: EcoR I, 5: Hind III and 6: BamH I. The sizes of the labelled DNA fragments are indicated. Figure 2B: genomic organization of the Leishmania (V) panamensis KMP-11 gene locus. B: schematic representation of the possible genomic organization of the KMP-11 locus. Shaded boxes indicate the position of the coding regions of the KMP-11 locus. The restriction sites and length of DNA fragments are indicated.
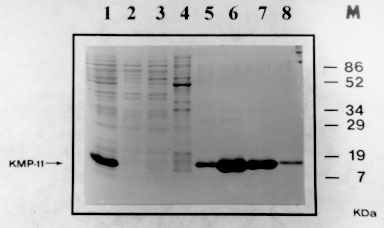
A: SDS-PAGE analysis of the expression and purification of the His-tagged recombinant KMP-11 protein. At each purification step, an aliquot was electrophoresed and the gel stained with coomasie blue. 1: supernatant phase of sonicated bacterial culture, 2: column flowthrough, 3 and 4 : washing fractions, 5, 6, 7 and 8: elution fractions, M: molecular weight marker. Figure 4B: Escherichia coli expression and western blot analysis of Leishmania. (V) panamensis KMP-11 protein. B: western blot of KMP-11 using serum of two VL patients (1, 2), rabbit polyclonal serum raised against the L. (L) infantum rKMP-11 (3), L98 mAb (4) and negative control (no primary antibody was added) (5). The immunoreactive band corresponding to a 14 kDa protein is indicated.
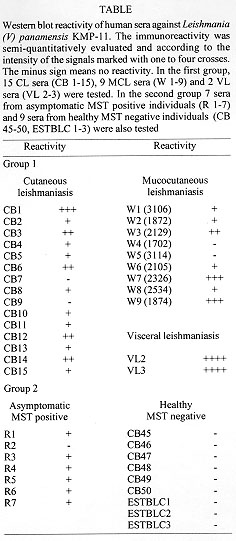
Antigenicity - The ability of the KMP-11 protein to induce proliferation of primed murine and human T-lymphocytes has been widely documented (Handman & Mitchell 1985, McConville et al. 1987, Russell & Alexander 1988, Jaffe et al. 1990, Jardim et al. 1991, Mendoca et al. 1991, Tolson et al. 1994). The B-cell antigenic potential has been also recently reported in dogs naturally infected with L. (L) infantum (Berberich et al. 1997). To date, a systematic study about the presence of specific anti-KMP-11 anti-bodies in humans suffering from different forms of leishmaniasis has not been carried out. Therefore, in order to analyze the antigenicity of KMP-11 during New World Leishmania infections in humans, the recombinant protein was assayed by western blots using sera obtained from CL, MCL and VL patients. In a similiar way, asymptomatic ("resistant") Leishmania-infected persons were tested. The Table shows that the rKMP-11 protein was recognized by 80% (12/15) and 77% (6/7) of sera from CL and MCL patients, respectively, infected with L. (V) panamensis. A high degree of variability in the levels of anti-KMP-11 antibodies was observed in these groups of patients. However, sera from several CL and MCL patients showed a very strong reactivity, evidencing that the protein may act as a potent human B-cell immunogen. Both (100%) of the human VL sera obtained from L. (L) chagasi-infected persons showed very strong immunoreactivity against the heterologous KMP-11 (Table) from L. (V) panamensis, consistent with the high levels of anti-Leishmania antibodies observed in these patients (Sacks et al. 1993). In addition, the immunological cross-reactivity between these VL sera and the L. (V) panamensis protein confirms the high level of conservation of KMP-11 between species causing either CL, MCL or VL. The potent B-cell immunogenic role of KMP-11 was also demonstrated when assaying sera from asymptomatic Leishmania-infected persons in western blots. A total of 86% (6/7) of these sera were seropositive for KMP-11, although, in general, the immunoreactivity was weak compared with the titers observed in most of the CL, MCL and VL sera. The low level of anti-Leishmania antibodies in these asymptomatic infections is consistent with the predominant Th1-like cellular immune reponse found in autohealing CL and patients without clinical symptoms of leishmaniasis (Carvalho et al. 1995). The negative control group (non-Leishmania infected persons) in contrast, did not show any reactivity against KMP-11 indicating lack of natural IgG antibodies against this protein. DISCUSSION Recent reports on the cloning and molecular characterization of the KMP-11 protein in the Old World Leishmania species L. (L) donovani and L. (L) infantum (Jardim et al. 1995b, Berberich et al. 1997) and in the Trypanosoma species T. b. rhodesiense (Stebeck et al. 1996) revealed similiar structural features of this kinetoplastid-specific protein, although differences in the genomic arrangement and in the pattern of distribution were found that may influence its function. The present study was carried out to analyze the sequence and gene locus organization of KMP-11 in the New World Leishmania species L. (V) panamensis, the prevalent etiological agent of cases of CL and MCL in Colombia (Corredor et al. 1990, Colombian Health Ministry 1994). Here, we provide evidence that the sequence of KMP-11 is highly conserved (> 95% homolgy) in all Leishmania species so far analyzed as well as the genomic organization consisting of three gene copies, regardless of the epidemiological pattern and the clinical symptoms of leishmaniasis they cause. The T. b. rhodesiense counterpart of this protein, in contrast, shows 18% divergence with respect to Leishmania and the gene is transcribed from a single gene copy (Stebeck et al. 1996). Immunofluorescence studies also showed that the L. (L) donovani (Stebeck et al. 1995), the L. (L) infantum (C Berberich, manuscript in preparation) and the T. b. rhodesiense (Stebeck et al. 1996) KMP-11 exhibit different fluorescence signals pointing to strict species-specific differences in the distribution of this protein. The biological function of this protein is still far from being clear. Recently, it has been speculated that the monomethylarginine in the L. (L) donovani KMP-11 sequence on position 45 might be involved in parasite-defense mechanisms (Jardim et al. 1995a) after partial degradation of the protein in the parasitophorus vacuole. This modified arginine residue is known as a competitive inhibitor of nitric oxide synthetase, a central enzyme involved in generation of the anti-Leishmania radical nitric oxide within activated murine macrophages (Liew et al. 1990). It was proposed that within the strongly hydrolytic conditions of the phagolysosome, the KMP-11 protein would be released from the parasite and degraded to amino acids by proteolytic enzymes. Subsequently the inhibitory monomethylarginine would contribute to the enzyme inhibition and parasite survival (Jardim et al. 1995a). However, in the L. (V) panamensis sequence as well in the L.(L) infantum (Berberich et al. 1997) and T. b. rhodesiense (Stebeck et al. 1996) molecules, this arginine was substituted by a lysine and should therefore not account for such a speculative function. Since the consistent co-purification of a minor component together with the full-length L. (L) donovani KMP-11 indicates a specific proteolytic event and, since major post-translational modifications including O-glycosylation and N-methylation of this protein seem to occur (Jardim et al. 1995a), we cannot, however, exclude that a monomethylarginine residue from another position might be responsible for such an effect. Both, the exact pattern of post-translational modifications and a potential inhibitory role of putative monomethylarginines in the KMP-11 molecule remain to be determined. So far, what can be assumed is, that the high level of sequence conservation of KMP-11 together with the wide distribution throughout kinetoplastid parasites, suggest an essential role of this protein in the parasite biology. Protein modeling analysis and detailed CD-experiments (Jardim et al. 1995b, M Fuertes & C Berberich, manuscript in preparation) suggest that KMP-11 has a highly amphipatic character with an unusually high degree of alpha-helical structure sharing important thermodynamic features with the class of apolipoproteins (Ryan et al. 1993). It has been therefore hypothezised, that the KMP-11 protein could be involved in the stabilization of the parasite membrane. However, recent findings of a strict developmentally regulated and stage-specific expression together with a defined cellular localization mainly around the flagellum and flagellar pocket in L. (L) infantum and T. b. rhodesiense (Stebeck et al. 1996, C Berberich, manuscript in preparation) suggests additional functions. The detailed antigenicity analysis of the recombinant KMP-11 of L. (V) panamensis in human CL, MCL and VL sera suggests that this protein acts as a potent B-cell immunogen during natural course of disease. Our results show that 82% (27/33) of Leishmania-infected humans (symptomatic or asymptomatic) contain detectable levels of anti-KMP-11 IgG antibodies ( Table). In general, a high degree of variability in the anti-KMP-11 antibody level was observed in CL and MCL sera and a stronger immunoreactivity is associated with symptomatic infections when compared with the asymptomatic form. Therefore, the high antigenicity of KMP-11 may point to a potential serological marker for diagnosis of infection and disease. However, in view of the immunological cross-reacitivity observed against the T. cruzi counterpart of KMP-11 (Stebeck et al. 1995) it may be necessary to determine specific epitopes in KMP-11 capable to distinguish between leishmaniasis and Chagas disease. Future studies will also concentrate on the T-cell antigenicity in all forms of human and mouse leishmaniasis in order to evaluate KMP-11 as a possible vaccine candidate. This work was supported, in part, by grants CIM9605 and CIM9750 from the Committee for the Development of Research (CODI) from the University of Antioquia and by the grant 1115-14436-95 from the Colombian Agency for the Development of Science and Technology (Colciencias). JR Ramirez is recipient of a PhD fellowship from Colciencias. A Jaramillo and ID Velez were recipients of investigator awards from Colciencias. REFERENCES African trypanosomes and is found in a wide variety of kinetolastid protozoan parasites. Mol Biochem Parasitol 71: 1-13
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98047c.jpg] [oc98047a.jpg] [oc98047f.jpg] [oc98047b.jpg] [oc98047g.jpg] [oc98047d.jpg] [oc98047e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}