 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Population Biology of Rhodnius domesticus Neiva & Pinto, 1923 (Hemiptera: Reduviidae) under Laboratory Conditions
Alessandra A Guarneri, Carlos J Carvalho Pinto, Chris J Schofield/*,
Departamento de Microbiologia e Parasitologia, Universidade Federal de
Santa Catarina, Caixa Postal 476, 88040-900 Florianopolis, SC, Brasil
*London School of Hygiene and Tropical Medicine, WC1 E7HT, Received 20 September 1997; Accepted 18 December 1997
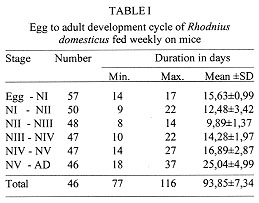
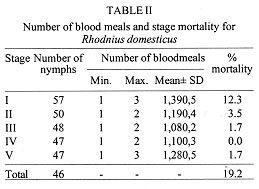
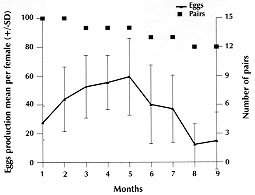
Code Number:OC98052 The entire life cycle of Rhodnius domesticus, fed weekly on mice, was studied under controlled conditions. Aspects related to hatching, life time, mortality, feeding behaviour and fecundity for each stage of the insect life-cycle were evaluated. The hatching rate observed in 100 eggs was 57% and the mean time of hatching was 15.6 days. Forty-six nymphs (80.7%) completed the cycle and the mean time from NI to adult was 93.8 days. The average span in days for each stage was 12.4 for NI, 9.8 for NII, 14.2 for NIII, 16.8 for NIV and 25.0 for NV. The number of bloodmeals in each nymphal stage varied from 1 to 3. The mortality rate was 12.3% for NI, 3.5% for NII and 1.7% for NIII and NV nymphs. The mean number of eggs laid per female in a 9-month period was 333.1. Average adult survival rates were 287.6 +133 and 328 +73 days for males and females respectively. Key words: Rhodnius domesticus - population biology - life cycle - triatomine The genus Rhodnius currently includes 13 species recognized on morphological characteristics (Lent & Wygodzinsky 1979, Lent et al. 1993). Of these, several are important as domestic vectors of Trypanosoma cruzi, the causative agent of Chagas disease, although the majority are of silvatic habit associated particularly with palm tree crowns (Schofield 1994). Some Rhodnius species are also vectors of T. rangeli, a parasite generally considered as being non-pathogenic to man (D'Ales-sandro-Bacigalupo & Saravia 1992). In spite of its name, Rhodnius domesticus Neiva & Pinto is known only from silvatic habitats in the Atlantic forest region of the states of Bahia, Espirito Santo, Rio de Janeiro, Sao Paulo, Parana and Santa Catarina. Little is known about the biological aspects of this species. However, it is found mainly in rodent or marsupial nests in epiphytic bromeliads, and occasionally in hollow trees. On the Island of Santa Catarina, R. domesticus populations from small mammal nests show infection rates with T. cruzi of over 50% (Galvao et al. 1961, Leal et al. 1961, Schlemper Jr et al. 1985). Experimental studies indicate that R. domesticus is also an important vector of T. rangeli amongst its mammalian hosts (Steindel et al. 1991). As part of a larger study on the comparative biosystematics of the genus Rhodnius, we present here the results of studies of the population biology of R. domesticus under experimental conditions. MATERIALS AND METHODS The insects used in this study were derived from a laboratory colony established in 1985 from bugs captured in the locality of Ribeirao da Ilha, Florianopolis, Santa Catarina. The colony is maintained at the Department of Microbiology and Parasitology of the Federal University of Santa Catarina where the bugs are kept at 25+/-4 C, 70+/-5% relative humidity (rh), and are fed weekly on immobilized white mice. Eggs from the colony were grouped by date of oviposition to initiate a cohort of 100 eggs. After eclosion, the first-instar nymphs were separated individually into plastic tubes (3.5 cm diameter x 4 cm height) lined with white filter paper and closed with cotton netting. Three days after eclosion, the nymphs were individually offered a feed on immobilized white mice during a 5 hr period each day until the first blood meal, after which they were offered a weekly feed. Bugs were maintained in a dark incubator at 28+/-1 C, 75+/-5% rh, and were checked daily for ecdysis or death. From the insects that completed development to the adult stage, 15 pairs were set up, each in individual tubes (8 cm diameter x 8 cm height) and maintained as before to determine adult survivorship and oviposition pattern. RESULTS The egg eclosion rate was 57% with a mean incubation period of 15.6 days (range 14-17 days). Forty-six nymphs completed the development to the adult stage (25 males and 21 females) (Table I), taking an average of 1.2 blood meals per nymphal stage (range 1-3) (Table II). The mean egg-to-adult development time was 93.8 days (range 77-116 days) (Table I). Adult survival ranged from 37 to 502 days for males (mean 287.6+/-133) and from 138 to 444 days for females (mean 328+/-73). The mean number of eggs laid per female was 335 (range 173-469) (Fig.).
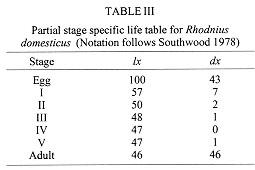
DISCUSSION The development cycle of Triatominae varies according to species and environmental conditions, and is greatly influenced by the availability of appropriate blood sources (Schofield 1985). In this study, rearing conditions were optimised as far as possible, with an absence of competition between individuals during their development. Under these conditions, development was rapid in comparison with other species of Rhodnius. Lent and Valderrama (1977) for example, found a mean development time of 114 days for R. prolixus, 181.2 days for R. neivai, and 190.7 days for R. pictipes, when the insects were fed weekly on restrained mice and maintained at 25+/-1 C and 70-75% rh. Under similar conditions, Jurberg and Rangel (1984) observed a mean development time of 358 days for R. pallescens. Soares et al. (1995) verified a mean development time of 209.4 days for R. nasutus fed weekly on chickens, and Rocha et al. (1994) found a mean development time of 278 days for R. pictipes fed fortnightly on pigeons and maintained at 28 C and 80% rh. Egg hatching rates in Rhodnius species also vary. They are generally above 80% (Szumlewicz 1975, Sherlock 1979) although lower average hatch rates of 53.3% and 64.9% have been recorded in established colonies of R. nasutus (Soares et al. 1995) and R. pictipes (Rocha et al. 1994) respectively. The 57% eclosion rate shown here may reflect the declining fertility of old females, since this variable was not controlled. This suspicion was confirmed in other experiments where we observed a hatch rate of over 80% in some egg batches from more recent colonies of R. domesticus (data not shown). Moreover, the rate of egg production in R. domesticus declines sharply from the fourth month after female emergence (Fig.) (cf. Guarneri et al. 1996). Mortality declined during the nymphal development stage, following a similar pattern to that observed in R. nasutus (Soares et al. 1995) and R. pictipes (Rocha et al. 1994). In the youngest nymphs, mortality appeared to be mainly due to an incapacity to feed, since dead bugs were generally without significant intestinal contents. In contrast, mortality in the older nymphs occurred mainly during their moulting. On average, about 80% of each stage required only a single meal in order to moult to the next. The short development time and high reproductive rate of R. domesticus recorded in this study probably represent close to the maximum achievable for this species under optimum conditions. The calculated rate of population increase (Ro = 70.4, r = 0.053) is considerably higher than that calculated for R. prolixus under broadly similar laboratory conditions (Ro = 49.3, r = 0.024) (Rabinovich 1972, 1974), and mainly reflects the much shorter generation time of R. domesticus. It is known, however, that the maximum potential reproductive rates are rarely achieved in nature, and it can be dramatically reduced in sub-optimal conditions. Studies of R. domesticus on the Island of Santa Catarina showed that this species is invariably present at very low population densities, and is found only in habitats formed by epiphytic bromeliads in association with the nests of small mammals (Leal et al. 1961, Schlemper Jr et al. 1985). In the course of our studies, we have found it impossible to maintain R. domesticus by feeding them on chickens, so it may be that in nature the population density is restricted primarily by limited access to mammalian blood sources. There have been marked environmental changes on the Island of Santa Catarina over the last few decades, which may have reduced the availability of mammalian hosts and bromeliad habitats, thus contributing to the scarcity of R. domesticus in this region. This study was partly supported by the ECLAT network, through EC grant number TS3*CT92-0092 and also by CNPq and CAPES. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98052b.jpg] [oc98052d.jpg] [oc98052a.jpg] [oc98052c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}