 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Ultrastructure of the Gastrodermis and the Nutrition of the Gill Parasitic Atriaster heterodus Lebedev and Paruchin, 1969 (Platyhelminthes: Monogenea) Claudia P Santos/^+, Thais Souto-Padron*,^ Reinalda M Lanfredi**
Universidade Santa Ursula, Instituto de Ciencias Biologicas e Ambientais,
Rua Fernando Ferrari 75, 22231-040 Rio de Janeiro, RJ, Brasil Received 19 November 1997; Accepted 8 January 1998
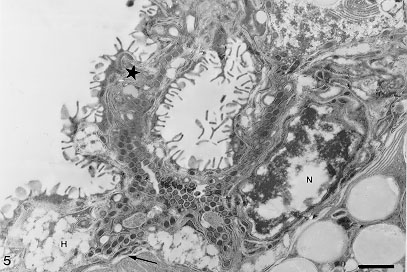
Code Number:OC98053 The gastrodermis of Atriaster heterodus Lebedev & Paruchin, 1969 (Polyopisthocotylea), a gill parasite from Diplodus argenteus (Valenciennes, 1830), is composed of "U"-shape hematin cells and a connecting syncytium, both having cytoplasmic lamellae. These cells show outgrowths and bent folds which were seen to enclose lumen material. The trapped material was then subjected to endocytosis. The nature of ingested food material was comparatively analyzed by cytochemical and histochemical tests. Blood residues were detected in the gut but tests for mucins were negative. No intact erythrocytes were observed in the gut lumen. Key words: Atriaster heterodus - Diplodus argenteus - Monogenea - fish - Brazil - nutrition The functional morphology of the digestive system has been studied for different Monogenea (Smyth & Halton 1983, Bogitsh 1993) and the feeding in polyopisthocotyleans and monopistho-cotyleans is generally distinct with the former ingesting blood (Llewelyn 1954, Halton 1982) and the latter, mucus and epithelial cells. However, blood pigments were observed in a few monopisthocotyleans (Uspenskaya 1962, Kearn 1963, Fournier 1978, Buchmann et al. 1987) while Allen and Tinsley (1989) demonstrated blood and epithelial cells at the same time in the gut lumen of a polyopisthocotylean species. Atriaster heterodus Lebedev & Paruchin, 1969 (Polyopisthocotylea) was recently reported from Rio de Janeiro coast (Santos et al. 1996). Surface topography and ultrastructural aspects of the spermatogenesis of this species have been described by Santos et al. (1997). The ultrastructure of the gastrodermis and the nutrition of this species are described herein. This paper represents the first ultrastructural study of gut caeca and nutrition of a marine monogenean from Brazil. MATERIALS AND METHODS Parasites - Sixty five specimens of A. heterodus were obtained from the gills of 25 Diplodus argenteus (Valenciennes, 1830) Sparidae, collected in Copacabana beach, Rio de Janeiro, Brazil. Histochemistry - Monogeneans and gill filaments were fixed in 5% buffered formalin or 70% ethanol. Samples were embedded in paraffin and cut in 5 mM sections which were deparaffinized with xylene and prior to staining, hydrated in decreasing concentrations of ethanol to water. Histochemical reactions are indicated with the references and color of positive reactions as follows. Neutral carbohydrate complexes: PAS (MacManus & Mowry 1960) (+=various shades of purplish-red); acidic carbohydrates: Alcian Blue (AB) and AB-PAS techniques (Mowry 1963) (+=blue); hemoglobin and derived pigments: Turnbull and Perls modified by Gomori (McManus & Mowry 1960) (+=blue) and after Lilly and Fullmer (1976) were tested Perls (+=blue-green), Okajima (+=orange-red) and Dunn-Thompson (+= emerald green). The picric alcohol solubility test (Llewelyn 1954) was tested for hematin (+=solve). Ultrastructure - For ultrastructural studies, parasites were fixed for 2 hr at 4 C in a solution containing 4% paraformaldehyde, 2.5% glutaraldehyde and 0.3M sucrose in 0.1M cacodylate buffer in sea water, pH 7.2. After being washed in the same solution, they were post-fixed for 1 hr in dark conditions in a solution containing 1% OsO4, 0.8% potassium ferricyanide, and 5mM CaCl2 in 0.1M cacodylate buffer, dehydrated in acetone, and embedded in Epon. Ultrathin sections were picked up on uncoated 200-mesh copper grids, stained with uranyl acetate and lead citrate, and observed in a Zeiss 900 or Jeol JEM 100CX Electron Microscope. Some ultrathin sections were subjected to the picric acid/alcohol solubility test for hematin (Halton et al. 1968). Control sections were incubated in acid-free alcohol. Cytochemistry - For basic protein detection, parasites were fixed as for ultrastructural studies, dehydrated in ethanol and incubated in a solution containing 2% phosphotungstic acid in absolute ethanol (E-PTA) for 2 hr at room temperature (Bloom & Aghajanian 1968). Specimens were rinsed in absolute ethanol, incubated for 10 min in propylene oxide and embedded in Epon to be observed with no counter stain. To test the presence of mucopolysacharides (Benhnke & Zelander 1970), parasites were fixed for 1 to 18 hr at room temperature in a solution containing 4% glutaraldehyde and 1% Alcian blue in sea water. Samples were washed twice for 10 min in sea water, and were post-fixed for 2 hr at room temperature in dark conditions in a solution containing 1% OsO4, 0.8% potassium ferricyanide and 5mM CaCl2 in 0.1M cacodylate buffer + 3% sucrose. Parasites were, then, dehydrated in acetone and embedded in Epon. Ultrathin sections were picked up on uncoated 200 mesh copper grids, stained with uranyl acetate and lead citrate. RESULTS The digestive caeca of A. heterodus are highly branched and interspersed within vitellaria. Their folded gastrodermis consists of isolated hematin cells in a connecting syncytium.
Hematin cells and syncytium show outgrowths and bent folds which were seen to enclose lumen material with the help of the surface lamellae. The trapped material was then subjected to endocytosis (Fig. 4). Residual pigments remain inside of the vesicles in the external region of the cell. The way they are released from the cell was not observed. No free hematin cell was observed in the gut lumen. The nature of hematin residues was confirmed by the histochemical test of picric acid in ethyl alcohol. The Turnbull method comparatively applied in the parasite and fish gill filaments confirmed the distribution of ferrous iron in hematin cells. The methods of Perls, Okajima and Dunn-Thompson were not conclusive. There was no reaction for mucin using the PAS and Alcian blue methods. Hematin cells stained with E-PTA displayed a strongly positive reaction in nuclear heterochromatin and in the small vesicles located at the external edge of the cell. Vesicles containing hematin residues were not labeled. A collection of membrane tubules and flattened sacs extends throughout the cytoplasm of the hematin cells. DISCUSSION The gastrodermis of A. heterodus is quite similar to the pattern described for other Polyo-pisthocotylea, with two cell types: hematin cell and connecting syncytium (Llewelyn 1954, Halton & Jennings 1965, Halton et al. 1968, Tinsley 1973, Halton 1975). Longitudinal sections showed that the irregular "U"-shaped hematin cells, are different from those previously reported for polyopisthocotyleans (Halton et al. 1968, Fournier 1978, Allen & Tinsley 1989). The feeding process is continuous, with lamellae looping back onto the surface to capture and endocyte food particles, in a process similar to that described by Halton (1974), actually called micropinocytosis. Fournier (1978) also mentioned the endocytosis of gut content but did not mention the importance of the lamellae in this process. The chemical nature of the pigments observed in the endocytosed vesicles was initially tested according to its solubility in picric acid in ethyl alcohol (Llewellyn 1954). Knowing that both formolic (formaldehyde) and hematin (hemoglobin) pigments are solved by picric acid and that formolic pigments do not react in the presence of Prussian blue (Perls), Turnbull blue, ferricyanide and ferrocyanide, contrary to what happen to hematin pigments (Lillie & Fullmer 1976), these tests were applied in samples fixed either in 70% ethanol and buffered formalin. As in sample fixed in 70% ethanol, where absence of formolic pigment is expected, the pigments were solubilized by the picric acid and also reacted for the Turnbull method, it was stated that the pigment was in fact hematin. The histochemistry and cytochemistry tests, negative for mucins and positive for iron residues, confirmed the chemical nature as hematin pigments, indicating hematophagous food habit for this parasite. Host hemoglobin in vacuoles strongly suggest that at least intracellular digestion occurs in these cells. No intact erythrocytes were observed in the gut. This can be explained by fish handling. The monogeneans were collected some hours after fish capture and according to Llewellyn (1954) erythrocytes can only be observed in their gut when fish are examined in less than one hour after capture, because a rapid haemolysis followed by phagocytosis normally occur. However, it cannot be excluded that erythrocytes are lysed during the feeding process. The structure of the syncytium cells with or without lamellae was previously reported (Halton et al. 1968, Tinsley 1973). Nevertheless, in both cases these monogeneans that feed on blood present a syncytium with few spread organelles, functioning mainly as a support for the hematin cells, on the contrary to those that present a mixed food habit like Polystomoides sp., where the syncytial cytoplasm has numerous organelles (Allen & Tinsley 1989). The reduced syncytial cytoplasm of A. heterodus with few organelles presupposes a support function but their role in the feeding process is still doubtful. This is the first study on the ultrastructure of the gastrodermis of A. heterodus with detection of its haematophagous food habit.
To Dr Kurt Buchmann (Royal Veterinary and Agriculture University, Denmark) and Dr Walter A Boeger (Universidade Federal do Parana, Brazil) who kindly commented on the manuscript. Financial support by CNPq (Conselho Nacional de Pesquisa) and PRONEX (Programa de Nucleos de ExcelEncia). REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98053b.jpg] [oc98053a.jpg] |
| |||||||||

{kind=link}
{kind=link}