 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Adhesion and Co-stimulatory Molecules in the Pathogenesis of Hepatic and Intestinal Schistosomiasis Mansoni Werner Jacobs^*, Eric Van Marck^+ Department of Pathology, University of Antwerp, Universiteitsplein 1, B-2610 Wilrijk, Belgium br
*Research Assistant of the F.W.O., Vlaanderen Received 17 March 1998; Accepted 7 May 1998
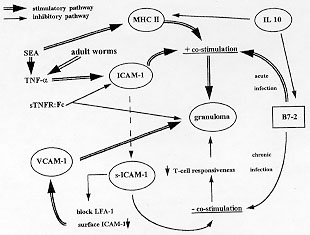
Code Number:OC98102 Infection of a susceptible host with the blood fluke Schistosoma mansoni results in the formation of periovular granulomas and subsequent fibrosis in the target organs. Granulomogenesis and fibrogenesis are mediated by immunological events which require cell-cell and cell-matrix interactions. In this review, the role of adhesion and co-stimulatory molecules in the genesis of the schistosomal pathology (granulomogenesis and fibrogenesis) is outlined. These molecules provide essential immunological interactions not only for the initiation of granuloma formation but also for the maintenance and modulation of the schistosomal granuloma during chronic infection. Furthermore, the role of secreted soluble adhesion molecules in the different clinical forms and in the modulation of the schistosomal granuloma is discussed. Recent new insights into the role of adhesion molecules for the induction of pathology by other developmental stages of the parasite (other than eggs) will be presented. Key words: adhesion molecules - co-stimulatory molecules - Schistosoma - pathogenesis - granuloma formation SCHISTOSOMIASIS Schistosomiasis is a helminthic disease, which causes considerable morbidity and mortality worldwide (WHO 1993). Infection of a susceptible host with the blood fluke Schistosoma mansoni results in the release of parasite eggs in the mesenteric and portal venous system, leading to entrapment of these eggs in the perisinusoidal spaces of the liver and in the intestinal wall. Tissue localization of antigen-secreting eggs initiates a persistent immunological insult to which the host responds by marshalling leukocyte recruitment and activation of inflammatory and immune-mediated responses resulting in granuloma formation and subsequent fibrosis. In a subpopulation of infected individuals, chronic granulomatous inflammation and fibrosis may cause hepato-splenomegaly, portal hypertension, and the development of oesophageal varices that may rupture and cause fatal bleeding, this being the main cause of death in this disease. During chronic infection, schistosome granulomas become modulated resulting in smaller hepatic and colonic granulomas (Colley 1975). Clinically, chronic S. mansoni infection in humans presents as a spectrum of disease forms, ranging from the relatively mild intestinal through the hepato-intestinal form and at the other end of the spectrum the severe hepatosplenic disease. The murine model of experimental schistosomiasis has been widely studied because it provides a unique model for the study towards the pathogenetic mechanisms underlying both granulomogenesis and fibrogenesis. The granulomogenic process in schistosomiasis is dependent on CD4+ T helper (Th) lymphocytes (Wynn & Cheever 1995) and results in a shift of the immunologic balance from a Th1 to a Th2 cell type response (Pearce et al. 1991, Grzych et al. 1991). ADHESION AND CO-STIMULATORY MOLECULES The emigration of circulating leukocytes into tissues and migration towards an antigenic nidus is a characteristic of the inflammatory response. Extravasation of leukocytes from the blood stream is controlled by the expression of cell surface adhesion molecules on both the circulating cells and on the vascular endothelium. Further immune responsiveness of T-cells to antigens and the effector activities of leukocytes requires cell surface molecules mediating cell-cell and cell-matrix interactions and providing additional co-stimulatory activity. Cellular adhesion molecules are currently classified into three families. In the immunoglobulin superfamily, the cell adhesion molecules are built up of immunoglobulin-like domains. Attention has been drawn to a very important feature of the immunoglobulin domain structure, namely mutual complementarity which allows strong interdomain noncovalent interactions. Major histocompatibility complex (MHC) molecules, the adhesion molecules ICAM-1, VCAM-1, and the co-stimulatory molecules B7/1 and B7/2 are some of the important members of this superfamily (Springer 1990, Freeman et al. 1993). The integrin family of adhesion molecules represents a group of proteins composed of noncovalently associated alpha and beta subunits. Integrins have been organized into three subfamilies based on the beta subunit utilization. The VLA (very late antigen) molecules (1 to 6) share a common beta1 subunit, but different alpha (1 to 6) subunits. VLA molecules act as receptors that bind to extracellular matrix components (Springer 1990). VLA-4 functions both as a matrix (fibronectin) and cell (VCAM-1) receptor (Osborn et al. 1989). The LFA-1 molecule, ligand of ICAM-1, belongs to the beta2 subfamily of integrins and is expressed on leukocytes. Integrins regulate leukocyte adhesion to endothelium, but where selectins, a family of Ca^2+ -dependent carbohydrate-binding proteins which exert functions in fast inducible intravascular adhesion and homing of lymphocytes, only mediate adhesion, integrins are still required for subsequent transendothelial migration (Johnston et al. 1989). Selectin adhesion receptors are molecules which have N-terminal lectin-like domains, an epidermal-growth factor-like domain and varying numbers of short consensus repeats in the extracellular part of the molecule (Springer 1990, Springer & Lasky 1991). Three selectins playing important roles in inflammatory events have so far been identified; E-, P-, and L-selectin. Selectins mediate adhesion of platelets to neutrophils and monocytes (P-selectin) (Larsen et al. 1990) or function in an early step of leukocyte binding to the endothelium, before transendothelial migration (L- and E-selectin) (Buhrer et al. 1990, Springer 1990), or mediate adhesion of lymphocytes to the specialized post-capillary endothelium of lymph nodes (L-selectin) (Camerini et al. 1989). LewisX (CD15) was identified as the selectin ligand (Larsen et al. 1990, Springer & Lasky 1991). SOLUBLE ADHESION MOLECULES IN SCHISTOSOMIASIS PATIENTS Recently, soluble forms of adhesion molecules were described in human serum. One of these soluble adhesion molecules, soluble ICAM-1, has a structure consistent with a membrane-cleaved variant of cellular ICAM-1 and maintains it ligand-binding capacity (Rothlein et al. 1991, Pigott et al. 1992). Soluble forms of ICAM-1 and E-selectin were found in the serum of patients with schistosomiasis mansoni (Secor et al. 1994, Lejoly-Boisseau et al. 1996). Adhesion molecules can be shed by activated endothelial cells (Pigott et al. 1992, Leeuwenberg et al. 1992). Serum levels of soluble adhesion molecules in schistosomiasis patients were significantly higher when compared to control patients and differences in the levels of soluble adhesion molecules were observed in patients with different clinical forms of schistosomiasis (Secor et al. 1994). Patients with high egg-secretion or hepatosplenic disease had significantly higher levels of s(oluble)ICAM-1 than individuals with low egg-secretion or intestinal disease. However, patients with intestinal disease had significantly higher levels of sE-selectin in their sera than did patients with hepatosplenic disease. No correlation could be found between the level of sICAM-1 and the schistosomiasis serological titre (Lejoly-Boisseau et al. 1996). Likely, higher levels of sICAM-1 reflect more intense inflammation in certain clinical forms, similarly as the correlation seen with severity in patients with primary biliary cirrhosis (Thomson et al. 1993). Malaria, often present in schistosomiasis endemic regions is a well know cause of elevated levels of sICAM-1 (Jakobsen et al. 1994). However patients infected with schistosomiasis, living in a malaria non-endemic zone also have higher levels of sICAM-1 when compared with healthy individuals (Lejoly-Boisseau et al. 1996). ADHESION MOLECULES IN SCHISTOSOMAL GRANULOMOGENESIS Granulomatous inflammation is one of the most complex cellular immune responses requiring recruitment and activation of a variety of bloodstream cells including monocytes, lymphocytes, neutrophils, and eosinophilic granulocytes. Cell adhesion between endothelial cells and leukocytes and between leukocytes mutually is a key event in the orchestration of cellular interactions which lead to the development of a circumscribed inflammatory infiltrate know as a granuloma. The adhesion molecules ICAM-1 and VCAM-1 are both expressed in S. mansoni egg granulomas (Rathore et al. 1996, Jacobs et al. 1997b,c), an observation consistent with the data of Lukacs et al. (1994), who could demonstrate an increase of ICAM-1 mRNA when eggs were injected in the lungs of mice to induce granulomas. Passive immunization of mice with anti-ICAM-1 monoclonal antibodies resulted in attenuation of granuloma development and reduced the proliferative response of spleen lymphocytes towards soluble egg antigens. However, whereas ICAM-1 is constitutively expressed in normal liver tissue, VCAM-1 is not (Rathore et al. 1996); VCAM-1 is dramatically induced with the onset of the hepatic egg deposition. Strong up-regulation of ICAM-1, its ligand molecule LFA-1, and the beta integrin VLA-4, but not VLA-6 are seen in schistosome granulomas (Langley & Boros 1995, Jacobs et al. 1997b,c). Interaction between ICAM-1 and LFA-1 appears to be the predominant interaction in schistosome egg granuloma formation, but when the ICAM-1 gene is knocked-out, up-regulation of VCAM-1 is seen and granuloma formation is preserved (Ritter & McKerrow 1996). VCAM-1 mediates adhesion of mononuclear to endothelial cells via binding to VLA-4 (Henseleit et al. 1994). Blockade of ICAM-1, LFA-1, and VLA-4 by monoclonal antibodies markedly inhibits spleen and granuloma cell interleukin (IL)2 and IL4 production - both cytokines being crucial for the initial events of schistosomal granuloma formation (Lukacs & Boros 1993, Wynn et al. 1993) - as well as lymphoproliferative responses, both in the acute and chronic infection (Langley & Boros 1995). As infection ages, granulomas appear with a non-homogenous ICAM-1/LFA-1 staining pattern, in contrast to homogenous staining granulomas during acute infection (Jacobs et al. 1997b). Whether this is due to the appearance of new cellular granuloma constituents that lack ICAM-1 or LFA-1 expression or that cellular expression is switched off in aging granulomas of previously immunoreactive cells is unclear, but since Langley and Boros (1995) did not observe apparent differences in the levels of expression of ICAM-1 and LFA-1 between acute- and chronic-infection granuloma derived cells, the appearance of new non-ICAM-1/LFA-1 immunoreactive cells in the granuloma co-existing with resident immunoreactive granuloma cells is most likely. It is remarkable that hepatocytes become ICAM-1 positive in some forms of hepatic inflammatory damage such as alcoholic hepatitis (Burra et al.1992), acute viral hepatitis B (Volpes et al. 1990), and liver allograft rejection (Adams et al. 1989), but not in acute or chronic hepatic schistosomiasis (Jacobs et al. 1997a,b). We could not detect hepatocyte ICAM-1 reactivity by immunohistochemistry nor by confocal microscopy. Using immunogold electron microscopy we were only able to demonstrate the presence of ICAM-1 on the microvilli of the hepatocyte cell membrane facing Disse's space (Jacobs et al. 1997b). Noteworthy is that all of these pathologies - except schistosomiasis - are the result of an immune aggression against the hepatocyte. ICAM-1-positive hepatocytes may thus be involved in facilitating the movements and possibly the guiding of cytotoxic lymphocytes towards hepatocytes (Volpes et al. 1990). Hepatic failure in schistosomiasis results from progressing liver fibrosis rather than immune-mediated hepatocyte damage. Furthermore, it should also be noted that histologically hepatocytes are in close relationship with ICAM-1 immunoreactive sinusoidal lining cells. Activated (sinusoidal) endothelial cells can shed ICAM-1 molecules as was demonstrated by Leeuwenberg et al. (1992). Secor et al. (1994) demonstrated elevated levels of soluble ICAM-1 molecules in the serum of schistosomiasis patients. ICAM-1 molecules thus might be released along the luminal side of the sinusoidal lining cells into the bloodstream where they can be detected as soluble ICAM-1 molecules. When they are released into Disse's space, mechanical trapping in the hepatocyte microvilli may occur (Jacobs et al. 1997b). We also observed upregulation of ICAM-1, LFA-1, and VLA-4 expression in ileal and colonic granulomas of S. mansoni infected mice (Jacobs et al. 1998). Morphologically, these observations could not explain the differences in granuloma size and down-modulation during chronic infection observed in these organs. The same set of adhesion molecules were also involved in the formation of hepatic (murine) S. haematobium granulomas (Jacobs et al., unpublished results). In addition, we could observe that S. mansoni worm-derived antigens induced ICAM-1/LFA-1-mediated granuloma formation in vivo (Jacobs et al. 1997a). Increased and early tissue expression of ICAM-1 was also seen in the skin of mice multiply infected with S. mansoni or vaccinated with gamma-irradiated cercariae after cercarial challenge (Ramaswani et al. 1997). Antigens derived from different parasite-stages or different schistosome parasites thus are capable of inducing increased localized expression of adhesion molecules in various organs of the host. S. mansoni granuloma T-cells express the same set of adhesion molecules (L-selectin^low, CD44^high, ICAM-1, LFA-1, and VLA-4) as granuloma T-lymphocytes in Mycobacterium avium infection (Sacco et al. 1996). Thus both Th1 (M. avium) and Th2 (S. mansoni) immune responses can give rise to granuloma infiltrating T-cells with the same adhesion molecule phenotype. This suggests that the mechanism for targeting immune cells to inflammatory sites is common to a wide variety of antigens. Schistosomal granuloma T-cells do not express the same set of adhesion molecules as splenic T-cells from S. mansoni infected animals (Sandor et al. 1995, Sacco et al. 1996). Their phenotype suggests that granuloma T-cells are almost exclusively memory/effector T-cells. Increased expression of ICAM-1 is preceded by the production of the pro-inflammatory cytokine TNF (tumour necrosis factor)-alpha. TNF-alpha has been identified as an important mediator of granuloma formation since it restores schistosomal granuloma formation in otherwise anergic SCID mice (Amiri et al. 1994). Lukacs et al. (1994) demonstrated that sTNFR:Fc constructs, which are effective in inhibiting TNF-mediated events, diminished schistosome granuloma formation and lymphocyte activation in vivo. Reduced schistosome granuloma formation was also seen in mice injected with anti-TNF antibodies (Joseph & Boros 1993). One of the mechanisms of TNF-alpha involvement in delayed type hypersensitivity schistosome granuloma formation is by up-regulation of TNF-alpha inducible ICAM-1 expression (Lukacs et al. 1994), which in turn perpetuates the antigen-specific response. APCs (antigen presenting cells), in addition to presenting processed antigen, also provide accessory cell-bound co-stimulatory signals that are necessary for stimulating maximal lymphokine production by CD4+ T cells. Co-stimulation is provided by a number of cell surface bound molecules such as MHC molecules, ICAM-1, and B7 molecules. Recent observations demonstrated that cells expressing the co-stimulatory molecule MHC class II are present in acute and chronic granulomas (Rathore et al. 1996) and that egg antigen-induced granuloma formation will fail to develop in MHC class II knock-out mice, but not in MHC I deficient mice because MHC class II deficient mice were unable to react to schistosome egg (Hernandez et al. 1997). These observations demonstrate that schistosomal granulomogenesis is mediated by MHC II binding CD4+ T helper cells and that MHC I binding CD8+ T cells play little role. Antibodies directed against B7-2 but not against B7-1 accessory molecules strongly inhibited egg-induced pulmonary granuloma formation and CD4 gene expression for IL4, 5, and 13 (Subramanian et al. 1997). Acute granulomas are rich in accessory (B7-2 antigen positive) cells that support helper T-cell activation, but as the infection becomes more chronic, more B7-2-poor cells appear in the granulomas and these cells are capable of inducing T-helper cell unresponsiveness (Rathore et al. 1996). Thus cellular interactions in acute granulomas may amplify egg-induced pathology, whereas in chronic granulomas they may precipitate down-modulation (Fig.). Increased expression of co-stimulatory B7-2 molecules is also observed in CBA/J mice displaying the more severe hepatosplenomegaly syndrome during chronic S. mansoni infection as compared to moderate splenomegaly syndrome mice (Freeman et al. 1996). Mice deficient for the B7/2 counter receptor generated an impaired Th2 cell response to S. mansoni infection (King et al. 1996). Downregulation of MHC class II and B7/2 molecules by IL10 results in downmodulation of egg antigen-specific delayed hypersensitivity and egg granuloma formation (Flores Villanueva et al. 1994, 1996) which induces a state of anergy that translates into the formation of downmodulated granulomas as typically observed during evolving schistosomal disease (Flores Villanueva et al. 1993).
The presence of lacto-N-fucopentaose III (LNFP-III), an oligosaccharide which contains the immunogenic LewisX trisaccharide was demonstrated on soluble egg antigens (Ko et al. 1990, Koster & Strand 1994). LewisX acts a ligand molecule for selectins (Larsen et al. 1990). LNFP-III induces proliferation of splenic B-lymphocytes and induces spleen B-cells of S. mansoni infected mice to produce IL-10 and PGE2 and thus down-modulate Th1 cells during experimental schistosomiasis (Velupillai & Harn 1994). Interaction between LNFP-III and B-lymphocytes could be mediated by a fucose-dependant ligand. One such receptor is P-selectin. Although P-selectin has not been found on B-cells (Springer & Lasky 1991), binding with a yet undescribed selectin is not excluded (Velupillai & Harn 1994). Alternatively LNFP-III may induce proliferation of activated B-cells through cross-linking with surface ICAM-1 on B-cells. This latter hypothesis is supported by the observation that LNFP-III is present on the a and b chain of LFA-1 (Forsythe et al. 1989), which is a ligand molecule for ICAM-1. Trottein et al. (1997) demonstrated that selectin carbohydrates were involved in antibody-dependent cell-mediated cytotoxicity of macrophages to schistosomula targets. Selectin-like counter receptors could be found on the surface of developing schistosomes. It is tempting to draw parallels between leukocyte-endothelium binding through adhesion molecules and the interaction between schistosome eggs with the vascular endothelium. Although there may be specific responses of the host endothelium to the S. mansoni egg, the endothelial egg interaction mediated by adhesion molecules appears to be generic until proven otherwise (File 1995). ADHESION MOLECULES AND GRANULOMA MODULATION Released soluble forms of adhesion molecules may have biological functions. Recombinant adhesion molecules have been demonstrated to block adhesion in vitro and in vivo (Watson et al. 1991). Locally high concentrations of shed adhesion molecules may thus impede adhesive interactions and provide a mechanism for negative feedback. L-selectin, but not of P- and E-selectin was observed immunohistochemically on the surface membranes of miracidia in tissue-trapped S. mansoni and S. japonicum eggs. El Ridi et al. (1996) suggested that circulating host-soluble L-selectin molecules, shed by L-selectin-positive leukocytes, transverse through the ultramicroscopic pores in the egg shell. These soluble L-selection molecules bind with a LewisX trisaccharide target ligand (Berg et al. 1992, Green et al. 1992). LewisX molecules are part of schistosome egg antigens (Ko et al. 1990, Koster & Strand 1994). The binding of sL-selectin to LewisX molecules in the schistosome egg may result in complex formation that ultimately impedes the release of soluble egg antigens and hence result in down-modulation of hepatic granuloma formation observed during chronic infection (El Ridi et al. 1996). An alternative explanation for the presence of L-selectin molecules on the miracidia may be that the parasite expresses host-like molecules, allowing it to escape the immune attack of the host. A striking feature is the inverse correlation between sICAM-1 levels in schistosomiasis patients and PBMC proliferative responses to egg antigens (Secor et al. 1994). Secor et al. (1994) formulated an interesting hypothesis linking sICAM-1 molecules with immune modulation of the hepatic schistosomal granuloma. Efficient activation of T-cells by APCs requires not only antigen presentation by APCs MHC class II molecules to the helper-T cell TCR, but also accessory signals mediated by co-stimulatory molecules and their ligands on APCs and T-helper cells. ICAM-1 plays an important role as a co-stimulatory molecule (van Seventer et al. 1990, Kuhlman et al. 1991). Therefore, if antigen presenting cells shed ICAM-1, they may not express sufficient levels of cell surface ICAM-1 for efficient co-stimulation of SEA-specific T-cells. Further, the shed ICAM-1 molecules can block proper co-stimulation by competitively binding to LFA-1 and thereby blocking the interaction of cellular ICAM-1 with its ligand. Either or both of these mechanisms would lead to reduced co-stimulation in the granuloma or draining lymph nodes and lead to inappropriate egg antigen presentation. Consequently, sICAM-1 released through the granuloma inflammatory response could provide a mechanism for feedback control of SEA-specific T-cell responsiveness, resulting in granuloma modulation (Fig.). Based on the currently available data it can be stated that in acute granuloma formation ICAM-1, LFA-1, VLA-4, and VCAM-1 are up-regulated, hence promoting cell-cell and cell-matrix interactions necessary for leukocyte activation and recruitment. The resulting juxtaposition enables interaction to occur between schistosome egg antigen laden macrophages (Bogers et al. 1996) bearing MHC class II molecules and SEA-specific CD4+ T-helper cells with the induction of IL2 and IL4 secretion. The presence of soluble ICAM-1 molecules, resulting in reduced levels of surface ICAM-1 and blocking of the LFA-1 ligand and the relative short-lived expression of B7-2 accessory molecules during acute infection may precipitate down-modulation of granuloma formation during chronic infection. This work was supported by grant 1.5.043.96 N of the Fonds voor Wetenschappelijk Onderzoek (F.W.O.), Vlaanderen. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98102a.jpg] |
| |||||||||

{kind=link}