 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Parasite Genotypically Related to a Monoxenous Trypanosomatid of Dog's Flea Causing Opportunistic Infection in an HIV Positive Patient Raquel S Pacheco /^+, Mauro CA Marzochi*, Marize Q Pires, Celia MM Brito*, Maria de Fatima Madeira*, Elizabeth GO Barbosa-Santos*
Departamento de Bioquimica e Biologia Molecular, Instituto Oswaldo Cruz,
Av. Brasil 4365, 21045-900 Received 16 January 1998; Accepted 29 May 1998
Code Number:OC98103 An HIV positive patient presenting a clinical picture of visceral leishmaniasis co-infection was submitted to a bone marrow aspiration after admission to hospital. Amastigotes forms were seen in the bone marrow aspirate and the parasite grew in culture as promastigotes. Molecular analyses showed that the flagellates isolated did not belong to the genera Leishmania, Trypanosoma or Sauroleishmania. It was not possible to establish infection in laboratory animals. In vitro culture of mouse peritoneal macrophages revealed the invasion of the host cells by the flagellates and their killing 48 hr after infection. Opportunistic infection with an insect trypanosomatid was suspected. Further hybridization analyses against a pannel of different monoxenous and heteroxenous trypanosomatids showed kDNA cross-homology with Leptomonas pulexsimulantis a trypanosomatid found in the dog's flea. Key words: opportunistic infection - AIDS - monoxenous trypanosomatid - minicircle - hybridization
The family Trypanosomatidae of the order Kinetoplastida is characterized by the presence of a kinetoplast-mitocondrial complex rich in DNA. This family includes parasite flagellates that undergo cyclical development in both vertebrate and invertebrate hosts. These parasites are best known as agents of important diseases in humans, domestic animals and plants. In addition to these heteroxenous organisms, several genera such as Crithidia, Herpetomonas, Blastocrithia and Leptomonas are restricted to a single host (monoxenous). Usually they are found infecting different insect orders such as Diptera, Hemiptera and Siphonaptera (Wallace 1966, Vickerman 1976, 1994). Such monoxenous or lower trypano-somatids have never been confirmed as pathogenic for human beings. Five species of the genus Leptomonas hosted by fleas (Siphonaptera) have been listed (McGhee & Cosgrove 1980). These parasites exist as attached and free promastigotes and amastigotes in the gut of the dog fleas. Amastigote forms are resistent to external enviroment and are deposited via feces of infected fleas (McGhee & Cosgrove 1980). More recently, Beard et al. (1989) reported the isolation of a new species of Leptomonas (Leptomonas pulex-simulantis, ATCC 50186) from the dog's flea Pulex simulans. Giemsa-stained preparations of the flea feces colected from the scalp of a dog demonstrated the presence of round aflagellate forms. In the past few years, new pathological aspects of protozoan infections caused mainly by Leishmania and Trypanosoma cruzi have emerged in immunosupressed patients. Particularly in cases of HIV co-infection the clinical symptoms are frequently peculiar. Cases of central nervous system tumor-like lesions (Ferreira et al. 1991, Pittella 1993, Rocha et al. 1994, Pacheco et al. 1998) and cutaneous lesions manifestations (Amato et al. 1997) related to the reactivation of Chagas' disease have been reported in association with immunosuppression. Distinct Leishmania isoenzymatic patterns and specific dermotropic zymodemes have also been found causing visceral leishmaniasis (Campino et al. 1994, Jimenez et al. 1995, Alvar et al. 1997). A recent publication reported (Dedet et al. 1995) a case of diffuse cutaneous infection caused by a presumed monoxenous trypanosomatid in a patient infected with HIV. The patient developed a diffuse leishmaniasis-like syndrome with numerous amastigotes in the skin nodules. The parasite isolated differed isoenzymatically from Leishmania, Trypanosoma and Sauroleishmania. Another interesting paper (Jimenez et al. 1996) reported the finding of an unusual Leishmania-like parasite in a case of visceral leishmaniasis/HIV co-infection. No kDNA sequence homology was observed with Leishmania nor with other genera of lower trypanosomatids in cross-hybridization experiments. The authors stressed that immunocompro-mised patients could be vulnerable to other non-human trypanosomatids. The present paper describes the isolation and the genotypic and phenotypic characterization of a monoxenous trypanosomatid found in the bone marrow of an HIV positive patient presenting a visceral leishmaniasis-like syndrome. MATERIALS AND METHODS Patient medical history - A 35-year-old heterosexual male, living in the district of Jacarepagua, Rio de Janeiro, Brazil was admitted to hospital in 1994 complaining of weight loss and weakness. The physical examination revealed enlargement of cervical lymph nodes, splenomegaly and fever. Laboratory analyses showed basically a hypochromic anemia and the hematocrit was 36%. The white blood cells and platelets count was normal. Serum chemistry values were within normal limits except for a serum alkaline phosphatase level of 9.0 U/100 ml [normal range 1.5 to 4 U (Bodansky)]. Serological test for hepatitis (A, B and C), toxoplasmosis and syphilis were negative. HIV infection was later diagnosed using enzyme-linked immunoabsorbent assay (ELISA) and Western blot techniques but unfortunately a CD4 count was not obtained. The patient presented tatoo marks on his body and denied belonging to the intravenous drug user group or to other risk groups. He is resident in an area considered as endemic for cutaneous leishmaniasis. Although no cutaneous scar had been found, the first clinical suspicion was that of visceralizing L. (Viannia) braziliensis infection. Immunofluorescence antibody test (IFAT) for leishmaniasis was high with a titer of 1/320. Search for amastigotes forms in the bone marrow aspirate was positive. The patient was treated for 20 days with N-methyl-glucantime antimoniate (20 mg/kg per day) and recovered completely. No recurrence was detected over a period of two years after treatment. Parasite isolation and stocks of trypanosomatids used as reference - A sample from the patient's bone marrow aspirate was inoculated in biphasic medium containing penicilin (1000 U/ml, Gibco) and streptomicin (1000 U/ml, Gibco). Parasites were subsequently propagated in Schneider's Drosophila medium (Gibco) supplemented with heat-inactivated 10% fetal bovine serum and 1% glutamine. The same culture was utilized to prepare the cell mass for phenotypic and genotypic analyses and cryopreserved in liquid nitrogen. Trypanosomatids used as reference in this study were as follow: L. (Viannia) braziliensis (MHOM/BR/75/M2903), L. (V) guyanensis (MHOM/BR/75/M4147), L. (Leishmania) amazonensis (IFLA/BR/67/PH8), L. (L) chagasi (MHOM/BR/74/PP75), S. tarentolae (ATCC 30267), T. cruzi (Y strain), Endotrypanum schaudinni (IM217), Phytomonas davidi (ATCC 30287), Blastocrithidia culicis (ATCC 30268), Crithidia deanei (ATCC 30255), Herpetomonas samuelpessoai (ATCC 30252), L. lactosovorans (ATCC 30970) and L. pulexsimulantis (ATCC 50186). Parasite study Mouse peritoneal macrophage culture - The methodology used has been previously reported (Eslami & Tanner 1994). The peritoneal cavity of outbread Swiss-Webster mice was each washed with 5 ml of RPMI medium (Sigma). The resident peritoneal cells were pooled, counted with a haemocytometer and made up to 2 x 10^6 viable cells/ml in RPMI medium. The cell suspension was placed onto several sterile coverslips within a culture flask and incubated at 37 C. Culture promastigotes were centrifuged prior to the inoculation at 900 G for 10 min at 4 C, counted and adjusted at a ratio of 1:5 (macrophage/parasite) in RPMI. After different times (day 0, 1, 2 and 5), the coverslips were fixed in methanol and stained with Giemsa in order to observe the parasites in the macrophages. Multilocus enzyme electrophoresis - Isoenzyme electrophoresis was performed according to Momen et al. (1985) and Pacheco et al. (1994) in 1% agarose gel using the following enzymes: aconitase (ACON, E.C.4.2.1.3), phosphoglucomutase (PGM, E.C.1.4.1.9), peptidase 3 (PEP-3, E.C.3.4.11), nucleotidase (NH, E.C.3.2.2.1), malate dehydrogenase (MDH, E.C.1.1.1.37), isocitrate dehydrogenase (IDH, E.C.1.1.1.42), peptidase D (PEP-D, E.C.3.4.13.9), peptidase 2 (PEP-2, E.C.3.4.11.1), malic enzyme (ME, E.C.1.1.1.40), glucose-6-phosphate isomerase (GPI, E.C.5.3.1.9), 6-phosphogluconate dehydrogenase (6PGD, E.C.1.1.1.43) and glucose-6-phosphate dehydrogenase (G6PD, E.C.1.1.1.49).
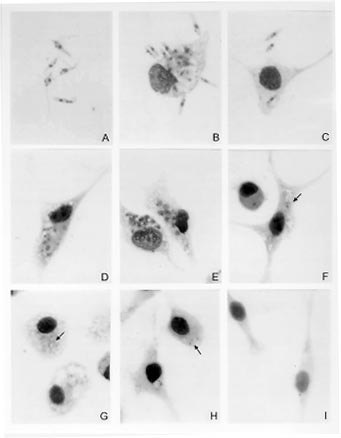
kDNA analysis and Southern blot hybridization - The technique of extraction and analysis of kDNA restriction profiles was carried out according to Pacheco et al. (1986). Purified kDNA preparations (1mg) were digested with the restriction enzymes Msp I and Hae III in the appropriate buffers according to the manufacturer's instruction. Fragments of maxi and minicircles were separated by electrophoresis in 1.5% agarose gels and stained by ethidium bromide. After electrophoresis, fragments were transferred to nylon membranes (Zetoprobe, Biorad) using the Southern's method (Southern 1975). After blotting the membranes were rinsed in 2 X SSC (300 mM sodium chloride/ 30 mM sodium citrate) and the DNA immobilized by ultraviolet light cross-link. Probes composed of cloned minicircles from L. (Viannia) braziliensis (MHOM/BR/75/M2903), L. (L) amazonensis (IFLA/BR/67/PH8), L. (L) chagasi (MHOM/BR/74/PP75) and total kDNA from the patient's isolate were radiolabelled with a ^32P dATP using the random primer method (KIT-Fiocruz, Brazil) to an specific activity of about 10^9 dpm/mg according to a modified protocol (Pacheco et al. 1994). Hybridizations were carried out at 65 C overnight, membranes were washed in 0.1X SSC/ 0.5% SDS at 65 C 3 times 30 min each and exposed to X-ray films overnight with an intensifying screen at -70 C. RESULTS Giensa-stained preparations of the parasite from Schneider's Drosophila medium showed pleomorphic flagellate forms with predominance of slender and shorter promastigotes. Round flagellate forms, although less frequent, were also observed. The size range of the promastigotes appeared to be larger than Leishmania promastigotes. Parasites grew profusely in the culture medium. Ultrastructure was examined by transmission and scanning electromicroscopy and excluded Trypanosoma infection (not shown). Macrophage culture - Peritoneal macrophages infected in vitro with promastigotes forms revealed internalized and degenerated parasites after 24 hr of infection. Nucleous and kinetoplast could hardly be observed. After 48 hr of infection, macrophages contained several vacuoles and small bodies. After five days, macrophages were found free of infection and presenting small inclusions, suggesting non-patogenicity or low virulence of the parasite (Fig. 1).
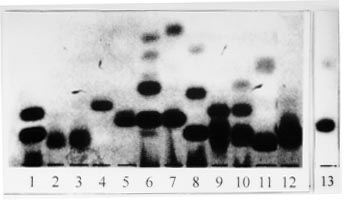
Isoenzyme electrophoresis - Results obtained using 12 enzymatic systems have allowed us to exclude any phenotypic identity of the isolate with Leishmania, Sauroleishmania, Trypanosoma, Endotrypanum, Phytomonas, Crithidia, Blastocrithidia, Leptomonas and Herpetomonas. Fig. 2 shows electromorphs for the enzyme MDH detected in the different genera.
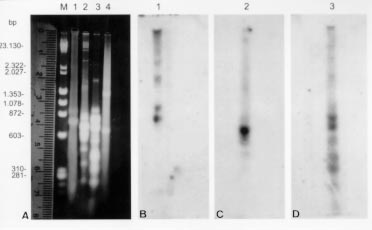
Genotypic analysis - The kDNA restriction profile of the isolate after digestion with the restriction endonuclease MspI showed no genotypic similarity with L. (V) braziliensis, L. (L) amazonensis and L. (L) chagasi (Fig. 3A). Southern blot hybridization of maxi and minicircles fragments with specific cloned minicircles of L. (V) braziliensis, L. (L) amazonensis and L. (L) chagasi used as probes revealed no sequence homology with the kDNA from the parasite isolated. Homologous sequences could only be detected with the respective homologous kDNAs (Fig. 3B, C, D).
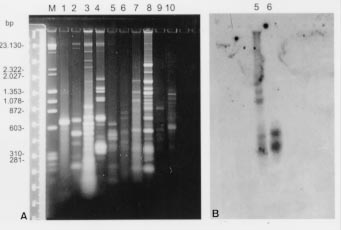
KDNAs from distinct species of trypano-somatids were further digested with the restriction enzyme HaeIII. Results showed in Fig. 4A revealed that each species displayed a different genotypic pattern. Nevertheless, when the radiolabelled kDNA from the human isolate was used as probe against the repertoire of trypanosomatids, homologous sequences in other minicircles fragments could only be found in the L. pulexsimulantis kDNA (Fig. 4B).
The presence of intracellular amastigote forms in Giemsa-stained preparations of bone marrow aspirates, the association with clinical and serological findings as well as the growth of the parasite predominantly as promastigotes have firstly led us to conclude that the patient was experiencing a visceral leishmaniasis/HIV co-infection. In immunocompetent individuals, even in spite of treatment and aparent cure of the disease, Leishmania parasites can persist and reactivate (Saravia et al. 1990, Guevara et al. 1993, Aebischer 1994, Schubach et al. 1998) raising the question on molecular determinants of virulence in some parasite strains. No relapse of the symptoms was observed in our immunocompromised patient over a period of two years. Evidence for low virulence of the parasite, in this case, was also supported by absence of cutaneous manifestations or visceral involvement after inoculation in the hind paws or via intraperitoneal in three golden hamsters. No histopathological alteration was observed in the skin, spleen and liver over a period of one year of observation (not shown). Likewise, reduced virulence was also evident from the parasite behavior within macrophages 48 hr after infections. The second step was to characterize the parasite isolated. Some areas in the district of Jacarepagua in Rio de Janeiro, where the patient is resident, are considered endemic for tegumentar leishmaniasis and cutaneous lesions are found in dogs and humans (Lopes et al. 1984, Pacheco et al. 1986). L. (V) braziliensis has been recorded ever since. In other foci near the metropolitan region of Rio de Janeiro both forms (tegumentar and/or visceral) of the disease are present in humans and domestic animals, being L. (V) braziliensis and L. (L) chagasi the unique species detected until now (Oliveira-Neto et al. 1986, Marzochi et al. 1994, Barbosa-Santos 1994). In such areas is not exceptional to find dogs and human beings living together in the same dwelling. Isoenzyme electrophoresis using 12 enzymatic loci and restriciton fragment length polymorphisms of kDNA analysis revealed patterns different from the most prevalent species of Leishmania causing tegumentar or visceral leishmaniasis in Brazil. Comparison with phenotypic and genotypic profiles of other genera of heteroxenous and monoxenous trypanosomatids also showed no identity. Hybridization of kDNA restriction fragments is frequently used to distinguish between strains and related species (Barker 1987). By using kDNA probes is possible to show that kDNA minicircles from organisms belonging to the principal Leishmania species complexes do not share extensive sequence homologies (Barker 1987). However, intraspecific variations are frequently detected using homologous kDNA probes (Pacheco et al., unpublished results). In the present case, no kDNA cross-homology could be detected between the parasite isolated and three different type-species representative of the Leishmania species complexes found in the New World. Although kDNAs from both human isolate and L. pulexsimulantis have displayed dissimilar restriction profiles, they share homologous sequences in minicircle fragments ranging from 603 to 310 bp. Whether the observed divergence is attributable to intraspecific variations or the in vivo generation of new polymorphisms under specific conditions (Pacheco et al. 1995) is difficult to ascertain. However, such homology seems to be specific and conserved in some minicircle classes in both kDNAs as no homologous sequences could be detected in minicircles from other species or even in another related Leptomonas species. This finding allows us to authenticate the close genotypic relationship between both parasites. The immune system depression in the patient might explain the opportunistic parasitism by this trypanosomatid. The presence of Leptomonas of the dog's flea in an HIV positive patient reinforces the idea that humans under immunossupression conditions may be vulnerable to other insect trypanosomatids giving rise to clinical manifestations similar to leishmaniasis. On the other hand it is not also so difficult to explain the mechanism of transmission being men and dogs living together. Feces deposited by infected adult fleas are usually well supplied with amastigotes which retain their infectiousness after drying at 25, 37 and 45 C (McGhee & Cosgrove 1980). The literature reported in 1980 a possible, whereas not proved, case of human infection by Herpetomonas (McGhee & Cosgrove 1980). The present article sustains that the possibility of lower trypanosomatids infecting humans exists and should be considered by attending physicians. ACKNOWLEDGMENTS To Prof. Maria Auxiliadora de Sousa (Nucleo de Tripanosomatideos, Instituto Oswaldo Cruz) for the donation of monoxenous trypanosomatids, Dr Maurilio J Soares (Departamento de Ultraestrutura e Biologia Celular, Instituto Oswaldo Cruz) for the electromicros-copy studies and Drs Claudio Siqueira, Tania Amaral and Luiz Franco (Setor de Hematologia do Hospital da Lagoa, Rio de Janeiro) for the hematological and clinical follow up of the patient. This work received financial support from the International Atomic Energy Agency (IAEA, Vienna, Austria), PAPES (Fiocruz) and Fundacao Nacional de Saude. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98103d.jpg] [oc98103a.jpg] [oc98103c.jpg] [oc98103b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}