 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
RESEARCH NOTE Massive Experimental Inoculation of Trypanosoma rangeli (Kinetoplastida, Trypanosomatidae) in Rhodnius neglectus Lent, 1954 (Hemiptera, Reduviidae) and Mus musculus Marina Vianna Braga/^+, Maria de Nazareth Meirelles*, Reginaldo Pecanha Brazil**
Laboratorio de Biologia e Controle de Insetos Vetores, Departamento de
Biologia Received 15 September 1997; Accepted 25 May 1998
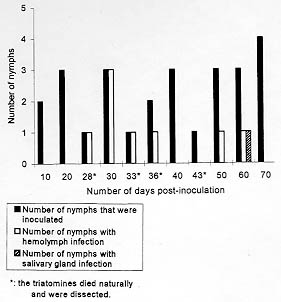
Code Number:OC98107 Key words: Rhodnius neglectus - Trypanosoma rangeli - Mus musculus - experimental infection RESEARCH NOTE The extra-intestinal development of Trypanosoma rangeli Tejera, 1920, in triatomines was studied by several authors and nowadays it is well known that this flagelate is able not only to develop in the hemolymph of the vector but also to invade its salivary glands being transmitted by the bite [R Zeledon 1965 Rev Biol Trop (Costa Rica) 13: 157-159, EJ Tobie 1965 J Parasitol 51: 837-841, MS Grewal 1970 Res Bull (NS) Panjab Univ 20: 449-480]. The infection can be transmitted both by the contaminative and inoculative methods, although this last one seems more efficient. The development of T. rangeli in the mammal host was accompanied in experimentally infected animals, specially rodents. In mice or rats, the parasites appear in the blood some hours after inoculation with trypomastigotes (from culture medium or salivary glands of triatomines), but in some cases only one or two weeks after, representing a period of incubation (H Groot 1952 Am J Trop Med Hyg 1: 585-592). The patent period in mammals is variable; in mice and rats experimentally infected it may last from three to seven months and eventually up to a year (EJ Tobie 1961 Exp Parasitol 11: 1-9). In humans with natural infection it persist from three to 18 months (F Pifano 1954 Arch Venez Patol Trop 2: 89-120). Rhodnius negectus Lent, 1954 were found naturally infected by T. rangeli in Brazil (L Diotaiuti et al. 1992 Mem Inst Oswaldo Cruz 87: 451). A study on experimental vectors of T. rangeli showed that among eight triatomine species tested, R. neglectus and Triatoma infestans (Klug, 1834) showed statistically higher rates of infection (A D'Alessandro 1972 J Med Ent 9: 187-195). Since it is well known that T. rangeli looses its capacity to develop in the insect and the vertebrate host after prolonged in vitro cultivation, the aim of our study was to verify the establishment of infection in R. neglectus by intra-celomical inoculation and in albino swiss mice, Mus musculus by intra-peritoneal inoculation with high concentrations of the Brazilian strain of T. rangeli, SC-58. R. neglectus were from a colony maintained at the Departamento de Biologia of the Instituto Oswaldo Cruz (IOC) obtained from the Laboratorio Nacional e Internacional de Referencia em Taxonomia de Triatomineos, Departamento de Entomologia, IOC. They were maintained at room temperature (22 C to 28.5 C and 70+/-5% of relative humidity). The 5th instar nymphs were separated from the colony five days after molting. The strain of T. rangeli used was SC-58, isolated by M Steindel et al. (1991 Mem Inst Oswaldo Cruz 86: 73-79) from a wild rodent, Echimys dasythrix (Hensel, 1872), by blood culture and by in vivo xenodiagnosis using nymphs of Panstronglylus megistus (Burmeister, 1835) and of T. infestans. The strain was mantained in biphasic culture medium, blood agar and LIT (liver infusion tryptose), supplemented with 10% of fetal bovine serum, at room temperature and the passages were made every two weeks. The triatomines had one leg cut off to obtain one drop of hemolymph and the abdomen compressed to obtain fresh posterior midgut contents. They were killed by freezing at 0 C, and the salivary glands were drawn out with tweezers and macerated. The midgut contents were also obtained by tearing the posterior midgut. The parasites, when found, were fixed with methanol (3 min), stained with Giemsa (10 min) and washed tap water. In the first experiment, the triatomines were inoculated intra-celomically with 0.1 ml of culture medium containing parasites: 5th instar nymphs (n=26) inoculated with 135.5 x 10^6 parasites per ml of culture medium and dissected on the 10th, 20th, 30th, 40th, 50th, 60th, and 70th day post-inoculation (p.i.). The triatomines were dissected also after the imaginal ecdysis. Those that died from natural causes were dissected on the 28th, 33rd, 36th, 43rd day p.i. After the inoculation, the triatomines were fed weekly on uninfected swiss mice until repletion. In the second experiment, two adult male swiss albino mice Balb-c (M. musculus) were inoculated intra-peritoneally with 1 ml of culture medium containing 62.0 x 10^6 parasites per ml. From the 3rd to the 60th day post-inoculation (p.i.), three 5th instar nymphs of R. neglectus were fed once, until repletion, on the swiss mice (3rd, 8th, 13th, 20th, 25th, 30th, 45th and 60th days p.i.) (a total of 24 nymphs) and dissected on the 15th days after the xenodiagnosis. One week after the infective meal, the nymphs were fed once on non-infected mice. Thirty one percent of the 26 inoculated triatomines were infected. On the 28th day p.i. the parasites were seen free in the hemolymph. On the 33rd day they were free and also seen adhered to the hemocytes. On the 36th day, one 5th instar nymph presented heavy hemolymph infection (more than 10 parasites per field), with some parasites showing signs of division. On the 50th day, only one 5th instar was infected, presenting a low number of parasites (less than 2 parasites per field). On the 60th day only female was infected (after imaginal ecdysis), and it was the only triatomine of this group to present salivary gland infection (extremely low number of parasites) (Fig. 1). None of them presented midgut infection.
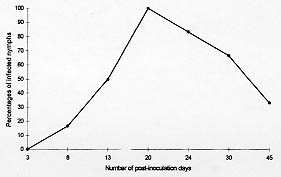
One hundred percent of the nymphs used for xenodiagnosis presented intestinal infection 15 days after feeding on mice on the 20th day after inoculation. Fig. 2 shows the percentages of infection of the nymphs used for xenodiagnosis. None of them presented hemolymph or salivary gland infection.
In our study, the 5th instar nymphs were inoculated with a high concentration of parasites per ml of culture medium. The percentage of triatomines that presented hemolymph infection was high, however, only one triatomine presented salivary gland infection (one female after the imaginal ecdysis), 32 days after the emergence of hemolymph infection. The number of parasites was usually low. Thus, our results suggest that the concentration of parasites in the inoculum does not have an influence on the establishment of the infection, only on its duration. JO Coutinho and V Nussenzweig (1952 Fol Clin Biol 18: 181-188) inoculated intra-celomically T. vitticeps, T. infestans, R. prolixus and P. megistus with one Venezuelan strain of T. rangeli and observed that the flagelates develop exclusively in the hemolymph when inoculated. MJ Soares (1982 J Parasitol 29: 529) inoculated intracelomically R. prolixus with a Colombian strain of T. rangeli, and observed free parasites only in the hemolymph. Our results are in accordance with these authors. G Rosa et al. (1995 Mem Inst Oswaldo Cruz 90 (Suppl): 299) made intra-femur inoculation of four triatomines species (including R. neglectus) with three strains of T. rangeli (including SC-58 from Brazil). The Brazilian strain presented low rates of hemolymph and salivary gland infection for R. neglectus (less than 10% of the inoculated triatomines). The Brazilian strain (SC-58) that was used by Rosa et al. (1995 loc. cit.), was the same that we used in our study, and their result for R. neglectus are different from ours since we obtained 31% of infection, probably due to the via of inoculation. Tobie (1961 loc. cit.) infected R. prolixus with several Venezuelan strains of T. rangeli by intra-celomic inoculation and observed that the flagelates were destroyed 24 hr after the invasion of the hemocoel, before they could reach the salivary glands. Zeledon (1965 loc. cit.) did not succeed in infecting R. prolixus by intra-celomic inoculation with a Panamanian strain. In our study, the parasites were not completely eliminated from the hemocoel of the inoculated triatomines, but, in most cases, they were unable to invade the salivary glands. According to Hoare (1972 loc. cit.), when the rodent was inoculated with infective forms isolated from salivary glands of triatomines, the parasites could be found in the blood from 10 to 14 hr after the inoculation. This author also reported that the blood parasitaemia, usually low, increased from the 4th day forward and could last months, but it could also become latent. Even though we used LIT as medium for the inoculum, it did not seem to have any influence on the growth of the parasites in the vertebrate host, since the parasitaemia of the mice inoculated was very low. EJ Tobie (1964 J Parasitol 50: 593-598) reported that two days after inoculation, the parasites could be found in the blood of the rodents. According to this author, it could be due to the strain of T. rangeli used (Venezuelan), and of the rodent. These results differ from ours since the swiss mice inoculated intra-peritoneally with T. rangeli, strain SC-58, developed latent infection, detected only by xenodiagnosis. Our results suggest that T. rangeli strain SC-58, may recover its infectivity after massive inoculation in the vertebrate host, although the infection was only detected in the blood of swiss mice by xenodiagnosis. It is also suggested that, even though R. neglectus is not considered the natural invertebrate host of the strain of T. rangeli used in these experiments despite the fact of having been found naturaly infected (Diotaiuti et al. 1992 loc. cit.), the infectivity of the strain SC-58 of the protozoa is partially recovered after massive inoculation in this species of triatomine, even though only one insect presented salivary gland infection. Further studies are needed to establish the number of passages through vertebrate and invertabrate hosts that are necessary for complete recovery of the infectivity of T. rangeli in the laboratory. Acknowledgements: to Dr A Romanha (Centro de Pesquisa Rene Rachou, Fiocruz, Brasil), Dr A Jansen (Departamento de Protozoologia, IOC) for providing the parasites, and AH Moraes Neto (Departamento de Biologia, IOC) for the critical review of the manuscript.
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98107b.jpg] [oc98107a.jpg] |
| |||||||||

{kind=link}
{kind=link}