 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 93 (5), September/October 1998, pp. 615-624 Molecular Epidemiology of Human Polyomavirus JC in the Biaka Pygmies and Bantu of Central Africa Sylvester C Chima+, Caroline F Ryschkewitsch, Gerald L Stoner Neurotoxicology Section, National Institutes of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, MD 20892, USA Received 15 June 1998; Accepted 30 July 1998
Code Number:OC98118
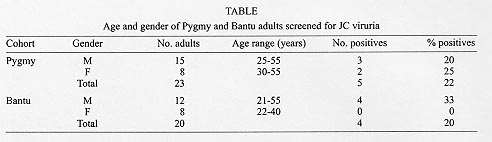
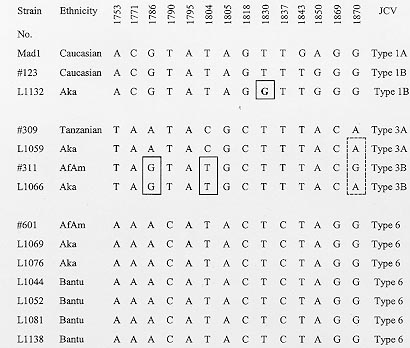
Polyomavirus JC (JCV) is ubiquitous in humans and causes a chronic demyelinating disease of the central nervous system , progressive multifocal leukoencephalopathy which is common in AIDS. JCV is excreted in urine of 30-70% of adults worldwide. Based on sequence analysis of JCV complete genomes or fragments thereof, JCV can be classified into geographically derived genotypes. Types 1 and 2 are of European and Asian origin respectively while Types 3 and 6 are African in origin. Type 4, a possible recombinant of European and African genotypes (1 and 3) is common in the USA. To delineate the JCV genotypes in an aboriginal African population, random urine samples were collected from the Biaka Pygmies and Bantu from the Central African Republic. There were 43 males and 25 females aged 4-55 years, with an average age of 26 years. After PCR amplification of JCV in urine, products were directly cycle sequenced. Five of 23 Pygmy adults (22%) and four of 20 Bantu adults (20%) were positive for JC viruria. DNA sequence analysis revealed JCV Type 3 (two), Type 6 (two) and one Type 1 variant in Biaka Pygmies. All the Bantu strains were Type 6. Type 3 and 6 strains of JCV are the predominant strains in central Africa. The presence of multiple subtypes of JCV in Biaka Pygmies may be a result of extensive interactions of Pygmies with their African tribal neighbors during their itinerant movements in the equatorial forest. Key words: polyomavirus - JC virus - genotypes - Pygmies - Bantu - Africa The dsDNA polyomavirus JC (JCV) is ubiquitous in humans and bears close sequence homology with other species of this genus, BK virus and the simian virus 40. Sero-epidemiologic studies have shown that up to 90% of adults are positive for antibodies to JCV (Walker & Frisque 1986). Infection with JCV is acquired in early childhood possibly via the respiratory tract. This is followed by persistent infection of the kidneys from which JCV is excreted in urine. Studies with polymerase chain reaction (PCR) show that 30-70% of adults worldwide are positive for JC viruria (Agostini et al. 1996, Sugimoto et al. 1997, Shah et al. 1998). JCV has been established as the causative agent in progressive multifocal leukoencephalopathy (PML), a fatal demyelinating disease of the central nervous system (Zurhein & Chou 1965). PML, previously a rare disorder found in immunocom-promised patients with hematologic malignancies, is now prevalent in 5-7% of AIDS cases in the USA and Europe (Berger & Concha 1995, Martinez et al. 1995), but in only 0.8% of Brazilian AIDS patients (Chimelli et al. 1992) and 1.5% in West African AIDS cases (Lucas et al. 1993). The complete genome of prototype JCV (Mad1) from the brain of a patient with PML was sequenced in 1984 (Frisque et al. 1984). The genome consists of a single molecule of dsDNA, 5.1kb in length, which is transcribed bidirectionally from the origin of DNA replication (ori). It codes for the early region proteins, large T and small t antigens which regulate transcription of the late region proteins VP1-3 and agnoprotein. JCV regulatory region can be classified into two major configurations: an "archetype" which is amplified from urine of normal individuals with JC viruria (Yogo et al. 1990) and a "PML type" when sequenced from the brain of patients with PML. PML-type regulatory regions are derived from the archetypal form by unique rearrangements, consisting of deletions and duplications within the JCV promoter/enhancer (Ault & Stoner 1993, Agostini et al. 1997c). Based on sequence analysis of JCV complete genomes, as well as segments of the VP1 and T antigen genes, JCV can be classified into several geographically based genotypes and subtypes (Ault & Stoner 1992, Agostini et al. 1995, 1997d, Sugimoto et al. 1997). The major genotypes so far described are Type 1, which is of European origin, Type 2, which is Asian, and Types 3 and 6 which are African in origin (Agostini et al. 1995, 1998). Type 4 which appears to be a recombinant of African and European Types (1 and 3)(Agostini et al. 1996), is prevalent within the United States with the highest frequency in African-Americans. A new clade of JCV strains, consisting of three possible subtypes has been identified in Southeast Asia (Ou et al. 1997) (Chima et al. unpublished data). Biaka Pygmies (singular `Aka'), are a group of aboriginal peoples in central Africa who live predominantly as hunter-gatherers in the tropical forest and have a shorter stature when compared to other Africans. Genetic studies have identified Pygmies to have distinctive genetic markers which may be described as "ultra-African" (Cavalli-Sforza 1986). The Biaka show a level of admixture with other Africans, with a residual incidence of 18-35% of ancient Pygmy genes (Cavalli-Sforza 1986, Cavalli-Sforza et al. 1994). It is estimated that the differences between Pygmies and their closest African neighbors are great enough to have required at least 10-20,000 years of isolation, considering that gene flow between this two groups occurs at the rate of only 0.7% per generation (Cavalli-Sforza 1986). The Biaka Pygmies presented in this study are members of the Babenzele clan, the easternmost subgroup of Aka or "Western" Pygmies, who live in the Dzangha-Sangha dense forest reserve on the banks of the Sangha river, below 4oN of the equator in Central African Republic (C.A.R) (Cavalli-Sforza 1986, Sarno 1995). The Bantu are African agriculturalists who speak a group of related languages and occupy the southern third of Africa starting from their putative origin in the Nigeria-Cameroon border in the west, to the Kenya-coastline in the east and as far south as Port Elizabeth in South Africa (Hrbek et al. 1992). Pygmies and their Bantu neighbors have a symbiotic relationship of mutual interdependence (Turnbull 1986, Bahuchet 1993, Sarno 1995). It is estimated that the Bantu first made contact with Pygmies during the Bantu expansion about 2-3,000 years ago (Cavalli-Sforza 1986, Hrbek et al. 1992). The Bantu villagers presented in this study live in close proximity and interact extensively with the Pygmies. Indeed, the Biaka and other Pygmy tribes speak a form of Bantu or Nilotic language borrowed from their neighbors having lost their own language over a long period of contact with other African tribes. However, ethnologists and linguists can still recognize common language elements between the Biaka in the west and the most genetically ancient and distant Pygmies (Mbuti), who live in the Ituri forest some 800 miles to the east (Bahuchet 1993, Sarno 1995). It is assumed that JCV, like any good parasite, has co-evolved with its human host. Due to the stable and distinct JCV genotypes which characterize different populations, urinary JCV has been shown to be a valuable tool in tracing human migrations (Agostini et al. 1997d, Sugimoto et al. 1997). To delineate the JCV genotypes circulating among the aboriginal peoples of central Africa, we undertook a study of the genotype profile of JCV excreted in the urine of the Biaka Pygmies and their Bantu neighbors with a view to determine whether unique strains of JCV may be circulating within these remote people and to compare the rates and pattern of JC viruria with other population groups around the world. MATERIALS AND METHODS Patients and samples - Single urine samples (5-50 ml), were collected from 33 Biaka Pygmies from the Pygmy settlement of Yandoumbe and 28 Bantu villagers from Amopolo within the Dsangha-Sangha dense forest reserve in Bayanga prefecture C.A.R. Seven additional urine samples were also collected from two female and five male Bantus living in the city of Bangui, C.A.R. There were 43 males and 25 females with an average age of 26 years and a range of 4-55 years. Adults 20 years and older made up 65% of the sample population. Age determination of the Pygmy population utilized educated estimates by an experienced Pygmy nurse practitioner. All subjects included in the study population were healthy volunteers. DNA extraction - Urine samples (5-15 ml) were centrifuged at 4,300 x g for 10 min and cell pellets were resuspended in phosphate buffered saline (PBS), recentrifuged and the supernatant was discarded. Cells were suspended in 100-200 ml digestion buffer containing 0.2 mg/ml of proteinase K, 50 mM KCl, 10 mM Tris/HCl (pH 8.3), 2.5 mM MgCl2, 10% (wt/vol) gelatin, 0.45% (vol/vol) NP40 and Tween20. After overnight incubation at 55oC in a waterbath, enzyme reactions were stopped by boiling for 10 min. DNA extracts were stored at -70oC until used and 2-10 ml of the extract was used for subsequent PCR. PCR - Initial tests for JCV were designed to amplify DNA fragments from the VP1 and large T antigen genes. JCV specific primers for the VP1 coding region were JLP-15 &16 which amplify a 215-bp fragment from this region. This DNA fragment provides up to 15 typing sites for differentiating JCV genotypes and subtypes (JLP-15, nucleotides 1710-1734, 5'ACAGTGTGGCCAGAATT CACTACC-3' and JLP-16, nucleotides 1924-1902, 5'-TAAAGCCT CCCCCCCAACAGAAA-3'). A segment of the large T antigen was amplified using the primer pair JTP-5&6 which amplify a 276-bp fragment from the T-antigen encoding the zinc-finger motif. This region is the site of a mutation changing a glutamine codon to leucine at amino acid 301. This point mutation is characteristic of all African and some Asian strains of JCV so far studied (Agostini et al. 1995, 1997a) (JTP-5 nucleotides, 3621-3642, 5'-CTTTGTTTGGCTGCTA CAGTAT-3' and JTP-6 nucleotides, 3896-3877, 5'-GCCTTAAGGAGC ATGACTTT-3'). The non coding regulatory regions and T-antigen intron were amplified using the primer pairs JRR-25 & 28 and JSP-1 & 2 respectively. JRR -25 & 28 amplify the entire regulatory region (341-bp) including three typing sites to the left of ori for distinguishing Types 1 and 2 strains (JRR-25, nucleotides, 4981-5004 5'-CATGGATTCCTCCCTA TTCAGCA-3' and JRR-28, nucleotides, 291-268 5'-TCACAGAAGCC TTACGTGACAGC-3'). Specific mutations at positions 133 and 217 of the archetypal regulatory region can be used to further characterize African genotypes. Deletion of certain pentanucleotide repeats within the regulatory region has been used to subtype JCV strains in Taiwan (Ou et al. 1997). The JCV specific primers JSP 1&2 amplify a 402-bp fragment from the T-antigen intron which provides additional typing sites for confirming genotype assignments (JSP-1 nucleotides, 4390-4412, 5'-ACCAGGATTCCCA CTCATCTGT-3' and JSP-2 nucleotides, 4791- 4769, 5'-GTTGCTCA TCAGCCTGATTTTG-3'). Following an initial heating at 94oC for 1.5 min (hot start), the 50-cycle, two-step PCR program include 1 min for annealing and elongation at 63oC, denaturation at 94oC for 1 min and extension at 72oC for 1 min. After a final extension for 10 min reactions were terminated at 4oC. PCRs were performed using UlTma DNA polymerase with 3'-5' proofreading activity (Perkin Elmer Cetus) in a standard buffer containing 1.5 mM MgCl2. Cycle sequencing - Gel-purified PCR products were sequenced directly using the Excel Kit (Epicentre Technologies, Madison, WI) with the same primers used for DNA amplification end-labeled with 33P-ATP (Amersham, Arlington Heights, IL). Initial denaturation at 95oC was followed by 30 cycles of 30 sec at 95oC for denaturation and 1 min at 63oC for annealing and elongation. Products were electrophoresed on a 6% polyacrylamide gel containing 50% urea. Gels were fixed with 12% methanol and 10% acetic acid, transferred to 3MM chromatography paper, dried under vacuum, then exposed to X-ray film for 12-48 hr. JCV genotypes were identified as previously described (Ault & Stoner 1992, Agostini et al. 1995, 1997b, 1997e, 1998). Sequence relationships were analyzed with GCG programs, Unix version 8 (Genetics Computer Group, Madison, WI). Primer design was assisted by the OLIGO program version 5.0 (NBI, Plymouth, MN). Reference sequences - The following are GenBank accession numbers for JCV sequences referred to in this work: JCV archetypal regulatory region JCV(CY) M35834 (Yogo et al. 1990); JCV coding region JCV(Mad-1), J02227 (Frisque et al. 1984); JCV Type 6 coding and regulatory regions, AF015537 and AF015538 (Agostini et al. 1998); JCV Type 3 strains #309, U73178, #311, U73501 (Agostini et al. 1997a); JCV strain#123, subtype 1B, AF015527 (Agostini et al. 1997b). RESULTS The age and gender of the Biaka and Bantu adults tested for JC viruria is given in the Table. Of the 43 adults tested by PCR amplification of the VP1 coding region, 22% (5 of 23) Pygmies and 20% (4 of 20) Bantus were shown to excrete the virus in urine. Overall, males had a higher excretion rate than females, seven out of 27 (26%) compared with two out of 16 (13%). None of the 24 children and adolescents aged 18 years or younger included in the sample population were positive for JC viruria. One of seven samples collected from Bantus in the city of Bangui was positive. This strain, L1081, was obtained from the urine of a 47-year old Cameroonian of the Bemoun tribe long domiciled in C.A.R. JCV coding regions - The JCV genotypes excreted by the nine adults were further analyzed by direct cycle sequencing of the JLP-15 & 16 amplified fragments from both directions. Within this fragment up to 18 typing sites have been identified for differentiating JCV genotypes and subtypes. Fourteen of these sites are illustrated in Fig. 1. JCV Type 6 can be clearly distinguished from both Types 1 and 3 at positions 1790 and 1837. Type 1 strains can be separated from both Types 3 and 6 at position 1771, while the two subtypes of Type 1, (1A and 1B) can be differentiated from each other at positions 1843 and 1850. Figure 1: typing sites within the JLP-15& 16 amplified fragments of the VP1 gene. Bantu and Pygmy strains are compared to JCV Mad1 sequence and strains #123 (Type 1B ) (Agostini et al. 1997b), #309 (Type 3A) from Tanzania, #311 (Type 3B) and # 601 (Type 6) from African-Americans (Agostini et al. 1997a, 1998). L1132 shows a point mutation at nucleotide 1830. L1066 shows similarity with Type 3B nucleotides at positions 1786 and 1804 (solid frame) , while it resembles Type 3A at position 1870 (broken frame). Numbering is based on the sequence of JCV Mad1 (Frisque et al. 1984). Analysis of the JCV strains from Pygmies yielded three different types of JCV from five positive samples. These were two Type 3 strains, one Type 1 and two Type 6 strains. One of the Type 3 strains (L1059) showed identical sequence in the VP1 fragment to the DNA sequence of strain #309 previously amplified from the urine of an African from Mara region in Tanzania (Agostini et al. 1995). The other Type 3 strain (L1066) showed partial sequence homology with #311(Type 3B), previously sequenced from an African-American, but differed from this strain at position 1870 where deoxyadenosine was inserted in place of deoxyguanosine. The latter strain was therefore termed a variant of Type 3B pending analysis of the complete genome. Strain L1132, from a Biaka Pygmy showed very close sequence homology in the VP1 fragment when compared to a Type 1B strain, #123, sequenced from a Caucasian (Agostini et al. 1997b). However this Aka strain had a distinct point mutation at position 1830, where deoxythymidine (T) was replaced by a `G'. This mutation caused a change in the codon for amino acid inserted at this position from valine to glycine. This point mutation at position 1830 of Aka strain L1132 has not been described previously in any Type 1 strains (Agostini et al. 1997b). Both Type 6 strains sequenced from Aka were identical with the previously reported Type 6 sequence (#601). A total of four JCV strains were sequenced from the Bantu. These four strains when analyzed showed exact sequence homology in the JLP-15 and 16 amplified fragments when compared to strain #601, sequenced from the brain of an African-American patient with PML. The Bantu Type 6 strains were also identical to the Aka Type 6 (Fig. 1). A 276-bp fragment was sequenced from the large T antigen of six JCV strains (three Aka and three Bantu) using the Primer pair JTP- 5 and 6. This T antigen fragment encodes the zinc finger motif. A specific point mutation in this fragment characterizes all African strains of JCV so far described and some Asian strains. This mutation is a non-conservative nucleotide base substitution at position 3768 from `T' to `A', causing a change in the amino acid coded from hydrophilic glutamine to hydrophobic leucine (Agostini et al. 1997a). The six Bantu and Pygmy strains amplified from the T-antigen zinc finger region showed a mutation at position 3768 (Fig. 2). Typing sites within this fragment confirm strain L1059 as a Type 3 strain and strains L1052, L1069, L1076, L1081 and L1138 as Type 6 strains. Figure 2: typing sites within the JTP-5&6 amplified fragment of large T antigen including the zinc finger motif. Position 3768 (frame) shows site of nucleotide mutation from "T" to "A" in all African genotypes including Bantu and Pygmy strains when compared to JCV Mad1. JCV noncoding regions - Noncoding regulatory regions of six JCV strains from Bantus and Pygmies were sequenced by the primers JRR-25 and 28 from both directions. The DNA sequence was compared to the consensus archetypal sequence of Type 1 (Agostini et al. 1996) and a Type 3 regulatory region sequence #309 from an Tanzanian (Agostini et al. 1997a). The Aka Type 3 strain (L1059) showed sequence identity with #309 including a point mutation at position 133 where `C' is characteristic of all Type 3 strains. Four Type 6 strains from Bantus and Pygmies, (L1052, L1069, L1076, and L1138) all showed an archetypal configuration without deletions. Strains L1081 (Type 6, Bantu) and L1059 (Type 3, Aka) both show a 10-bp deletion at nucleotides (51-60), just preceding the first NF1 site (Fig. 3). The deletion at this site is identical to those observed in strains #307 and #309 from Tanzania (Agostini et al. 1995, 1997a). All the Type 6 strains and the single Type 3 strain were characterized by the nucleotide "G" at position 217, however only the Type 3 strain showed deoxycytosine at position 133 of the regulatory region. Figure 3: regulatory region sequences amplified from Pygmy and Bantu strains is compared to the consensus archetypal regulatory region of Type 1 (Agostini et al. 1996) and #309 from Tanzania. Dashed lines denote uniformity with the consensus archetypal sequence. Solid lines show areas of nucleotide deletion initially observed in strains #307 and #309 (Agostini et al. 1995, 1997a) and now found in L1059 from a Biaka Pygmy and L1081 from a Bantu. At position 133, "A" is replaced by "C" in all Type 3 strains. At position 217, both Type 3 and Type 6 strains substitute deoxyguanosine for deoxyadenosine. Numbering is based on archetypal numbering of strain CY (Yogo et al. 1990). A 402-bp fragment was amplified from the noncoding T-antigen intron using the primers JSP-1 and 2. This fragment provides up to 15 additional typing sites for confirmation of JCV types and subtypes from the coding region sequences. Seven JCV strains were amplified from this fragment in the Pygmy and Bantu cohorts. Cycle sequencing confirmed the previous type assignments from the VP1 gene. L1044 (Bantu, Type 6) showed two nucleotide mutations at positions 4562 and 4648 while L1059 (Aka, Type 3) showed a single mutation at position 4435 (Fig. 4). The significance of these point mutations is unknown since the primary function of introns is to be spliced out prior to protein translation. Figure 4: the JSP-1&2 amplified fragment of the T antigen intron further confirm genotype assignments from the VP1 and large T antigen genes. Typing in this region is compared to the consensus sequence of Type 3 (Agostini et al. 1997a), strain #601 (Agostini et al. 1998) and Mad1. Framed sets denote sites of specific point mutations in L1044 and L1059 from Biaka Pygmies. Numbering is based on Mad1 sequence. DISCUSSION This study delineates the genotype profile of JCV strains circulating among the Biaka Pygmies and Bantu from Bayanga prefecture of C.A.R. This aboriginal African population excretes JCV in urine at a lower rate (21%) when compared to rates of excretion in urban populations in the United States (41%) (Agostini et al. 1996) and Europe (Stoner et al. 1998a). Native American tribes in the United States and the Pacific Islands show a rate of JC virus excretion in urine (65%) (Agostini et al. 1997d), which is three times the rate observed in this African cohort. However the rate of excretion among the Bantu and Pygmies are somewhat closer to a reported incidence rate of 30% in HIV positive patients from the Mara region of northwest Tanzania (Agostini et al. 1995). The reasons for the differences in rates of JCV virus excretion in different populations is not yet explained. However, it may be related in part to the difference in age of various sample populations. Studies in Caucasians and African-American cohorts within the United States have shown that the rate of JC virus excretion in urine rises dramatically in the fifth decade of life (Agostini et al. 1996), (Chima, unpublished observations). It therefore follows that sample populations with older age groups are more likely to yield a higher rate of JC viruria. The African cohort studied here had only three adults estimated to be aged 50 years or older. Analysis of the JCV strains from Pygmy urine revealed four different subtypes from the five positive cases. These were two Type 3 strains (one 3A and one 3B variant), two Type 6 and one Type 1B variant. The Type 3A strain showed close identity with Type 3 strains previously reported among Nilotic Africans of the Luo tribe from the Mara region of Tanzania. The Type 3B strain showed a similar sequence to that recently found in an African-American (strain A179) (Chima, unpublished data). This is a variant of strain #311 also found in an African-American with an `A' to `G' substitution at position 1870 of the VP1 gene. The two Type 6 strains were identical to those sequenced from the urine of the Bantu in this study. JCV Type 6 was first sequenced from the brain of an African-American patient with PML (Agostini et al. 1998). This was later identified as a new subtype of JCV when similar strains were sequenced from the urine of Africans from Ghana (Guo et al. 1996). Type 6 strains have also been sequenced from the brains of AIDS patients with PML from the Ivory Coast (Stoner et al. 1998b) as well as the urine of an immunocompetent individual from Sierra Leone (Chima, unpublished data). The four JCV strains excreted in the urine of Bantus reported here are Type 6. Of the four Bantu strains, (L1081) showed a 10-bp deletion in the regulatory region sequence similar to that found in #309 from Tanzania and L1059 in Pygmies. However, L1059 also displays another marker of Type 3 strains, i.e., deoxycytosine at position 133 of the archetypal regulatory region. It is more likely therefore, that these two strains arose independently of each other rather than as a result of viral recombination. We can hypothesize that the two African genotypes of JCV (Types 3 and 6) may have co-evolved, independently of each other, in their respective African hosts. All genotype studies on JCV in Africans so far have shown that both Type 3 and 6 strains can be found in West and Central Africa (Guo et al. 1996, Sugimoto et al. 1997, Stoner et al. 1998b), while Type 3 is the only genotype so far described from East Africa (Agostini et al. 1995). Archeological and linguistic data have shown that the Biaka Pygmies migrated to their present location from a region north of the Ituri around the southern Sudan, first to northern Zaire and then in a northwest direction to their present location in the southwest tip of C.A.R. around the Sangha river (Cavalli-Sforza 1986, Bahuchet 1993). The putative site of Biaka Pygmy origin around the southern Sudan is closer to the region occupied by previously studied Africans from northwest region of Tanzania. The latter population are in part Nilotics of the Luo tribe (Agostini et al. 1995). This group excrete Type 3 JCV strains similar to those found in Biaka Pygmies. The Bantus on the other hand are migratory farmers thought to have come into contact with the Pygmies about 2000 years ago during the Bantu expansion from West Africa (Cavalli-Sforza 1986, Hrbek et al. 1992). Archeologists and historians estimate that during the second stream of the Bantu expansion, there was a migration along the banks of the Sangha river into central Africa (Hrbek et al. 1992). It is therefore likely that Bantu descendants of the first immigrants still occupy the present location and carry JCV strains transmitted from their parents. Due to the close interaction between the Pygmies and their Bantu or Nilotic neighbors in equatorial Africa, it may be speculated that Type 6 strains were transmitted to the Biaka during their later interactions with Bantus while the Type 3 strains were brought along during their migration from southern Sudan and East Africa. A Type 1B variant of JCV was sequenced from the urine of a 55 year old female Pygmy. Type 1 strains are generally characteristic of Europeans. This Aka strain bears a unique mutation at position 1830 not previously reported in Type 1 strains of JCV (Agostini et al. 1997b, Stoner et al. 1998a). The significance of this Type 1 strain is unknown although in another study, it has been reported that a pocket of the European subtype of JCV was found in Bangui, C.A.R. (Sugimoto et al. 1997). Analysis of the complete genome of the Aka Type 1B variant and identification of more JCV strains with similar mutations will facilitate characterization of this subtype. It is possible that on analysis of the complete genome, this strain may represent a unique subtype of JCV different from Type 1 strains We conclude that human polyomavirus JCV is excreted in the urine of Biaka Pygmies and Bantus of central Africa, though at a lower rate than that observed in other population groups. This study confirms Types 3 and 6 as the predominant genotypes of JCV in central Africa. The finding of four different subtypes of JCV in the urine of Biaka Pygmies may be explained by the extensive interactions of Pygmies with their various African tribal neighbors over a long period of time, as they moved from place to place in the equatorial forest. ACKNOWLEDGMENTS To Hansjurgen T Agostini for initial studies on African genotypes of JC virus. To the entire staff of the World Wildlife Fund in Bangui and Bayanga for their kind hospitality and assistance throughout our stay in the Central African Republic. REFERENCES Agostini HT, Brubaker GR, Shao J, Levin A, Ryskewitsch CF, Blattner WA, Stoner GL 1995. BK virus and a new type of JC virus excreted by HIV-1 positive patients in rural Tanzania. Arch Virol 140: 1919-1934. Agostini HT, Ryschkewitsch CF, Stoner GL 1996. Genotype profile of humam polyomavirus JC excreted in urine of immunocompetent individuals. J Clin Microbiol 34: 159-164. Agostini HT, Ryschkewitsch CF, Stoner GL 1998 . The complete genome of JC Virus Type 6 from the brain of a African-American with progressive multifocal leukoencephalopathy (PML). J Hum Virol: in press. Agostini HT, Ryschkewitsch CF, Brubaker GR, Shao J, Stoner GL 1997a. Five complete genomes of JC virus Type 3 from Africans and African Americans Arch Virol 142: 637-655. Agostini HT, Ryschkewitsch CF, Singer CF, Stoner GL 1997b. JC virus Type 1 has multiple subtypes: three new complete genomes. J Gen Virol 79: 801-805. Agostini HT, Ryschkewitsch CF, Singer EJ, Stoner GL 1997c. JC virus regulatory region rearrangements and genotypes in progressive multifocal leukoencephalopathy: two independent aspects of virus variation. J Gen Virol 78: 659-664. Agostini HT, Ryschkewitsch CF, Yanagihara R, Davis V, Stoner GL 1997d. Asian genotypes of JC virus (JCV) in Native Americans and in a Pacific Island population: markers of human evolution and migration, Proc Natl Acad Sci US 94: 14542-14546. Agostini HT, Shishido Y, Ryschewitsch CF, Stoner GL 1997e. JC Virus Type 2: definition of subtypes based on analysis of ten complete genomes. J Gen Virol: in press. Ault GS, Stoner GL, 1992 . Two major types of JC virus defined in progressive multifocal leukoencephalopathy brain by early and late coding region DNA sequence J Gen Virol 73: 2669-2678. Ault GS, Stoner GL 1993. Human polyomavirus JC promoter/enhancer rearrangement patterns from progressive multifocal leukoencephalopathy brain are unique derivatives of a single archetypal structure. J Gen Virol 74: 1499-1507. Bahuchet S 1993. History of the inhabitants of the Central African Rain Forest: Perspectives from comparative linguistics, p. 37-54. In CM Hladik, A Hladik, O Linares, H Pagezy, A Semple & M Hadley (eds), Tropical Forests, People and Food, UNESCO, Paris. Berger JR, Concha M 1995. Progressive multifocal leukoencephalopathy: the evolution of a disease once considered rare. J Neurovirol 1: 5-18. Cavalli-Sforza LL 1986. African Pygmies, Academic Press, Orlando. Cavalli-Sforza LL, Menozzi P, Piazza A 1994. Africa, p. 159-194. In Cavalli-Sforza LL, Menozzi P, Piazza A (eds), The History and Geography of Human Genes, Princeton University Press, Princeton. Chimelli L, Rosemberg S, Hahn MD, Lopes MBS, Barretto-Netto M 1992. Pathology of the central nervous system in patients infected with the human immunodeficiency virus (HIV): a report of 252 autopsy cases from Brazil. Neuropathol Appl Neurobiol 18: 478-488. Frisque RJ, Bream GL, Cannella MT 1984. Human polyomavirus JC virus genome. J Virol 51: 458-469. Guo J, Kitamura T, Ebihara H, Sugimoto C, Kunitake T, Takehisa J, Na YQ, Al-Ahdal MN, Hallin A, Kawabe K, Taguchi F, Yogo Y 1996. Geographical distribution of the human polyomavirus JC virus type A and B and isolation of a new type from Ghana. J Gen Virol 77: 919-927. Lwango-Lunyiigo S, Vansina J 1998. The Bantu-speaking people and their expansion, p. 75-85. In I Hrbek, General History of Africa, Vol. 111, Africa from the Seventh to the Eleventh Century, UNESCO, Paris. Lucas SB, Hounnou A, Peacock C, Beaumel A, Djomand G, N'gbichi J-M, Yeboue K, Honde M, Diomande M, Giordano C, Doorly R, Brattegaard K, Kestens L, Smithwick R, Kadio A, Ezani N, Yapi A, De Cock KM 1993. The mortality and pathology of HIV infection in a West African city. AIDS 7: 1569-1579. Martinez AJ, Sell M, Mitrovics T, Stoltenburg-Didinger G, Inglesias-Rojas JR, Giraldo-Velasquez MA, Gosztonyi G, Schneider V, Cervos-Navarro J 1995. The neuropathology and epidemiology of AIDS. A Berlin experience. A review of 200 cases. Path Res Pract 191: 427-449. Ou W, Tsai R, Wang M, Fung C, Hseu T, Chang D, 1997. Genomic Cloning and sequence analysis of Taiwan-3 Human Polyomavirus JC Virus. J Formos Med Assoc 96: 511-516. Sarno L 1995. Bayaka: The Extraordinary Music of the Babenzele Pygmies Ellipsis Arts,New York, 66 pp. Shah KV, Daniel RW, Strickler HD, Goedert JJ 1998. Investigation of human urine for genomic sequences of the primate polyomaviruses simian virus 40, BK virus, and JC virus. J Infect Dis 176: 1618-1621. Stoner GL, Agostini HT, Ryschkewitsch CF, Komoly S 1998a. JC virus excreted by multiple sclerosis patients and paired controls from Hungary. Multiple Sclerosis 4: 45-48. Stoner GL, Agostini HT, Ryschewitsch CF, Mazlo M, Gullotta F, Wamukota W, Lucas S 1998b. Two cases of progressive multifocal leukoencephalopathy (PML) due to JC virus: detection of JCV Type 3 in a Gambian AIDS patient. J Med Microbiol 47: 1-10. Sugimoto C, Kitamura T, Guo J, Al-Ahdal MN, Schelnukov SN, Otova B, Ondrejka P, Chollet JY, El-Safi S, Ettayebi M, Gresenguet G, Kocagoz T, Chaiyarasamee S, Thant KZ, Thein S, Moe K, Kobayashi N, Taguchi F, Yogo Y 1997. Typing urinary JC virus DNA offers a novel means of tracing human migrations. Proc Natl Acad Sci USA 94: 9191-9196. Turnbull CM 1986. Survival factors among Mbuti and other hunters of the Equatorial Rain Forest, p. 103-123. In Cavalli-Sforza LL, African Pygmies, Academic Press, Orlando. Walker DL, Frisque RJ 1986. The biology and molecular biology of JC virus, p. 327-377. In NP Salzman The Papovaviridae, Vol. 1, The Polyomaviruses, Plenum Press, New York. Yogo Y, Kitamura T, Sugimoto C, Ueki T, Aso Y, Hara K, Taguchi F 1990. Isolation of a possible archetypal JC virus DNA sequence from non immmuno-compromised individuals. J Virol 64: 3139-3143. Zurhein GM, Chou SM 1965. Particles resembling papova viruses in human cerebral demyelinating disease. Science 148: 1477-1479. Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98118c.jpg] [oc98118b.jpg] [oc98118d.jpg] [oc98118e.jpg] [oc98118a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}