 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
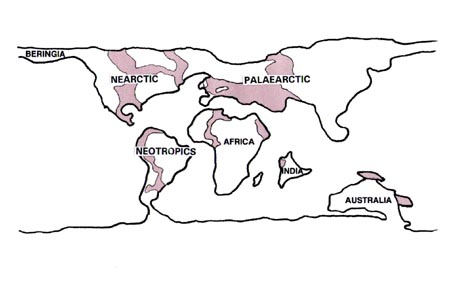
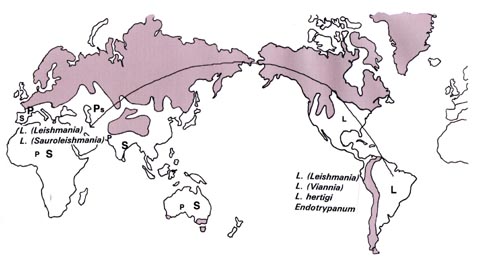
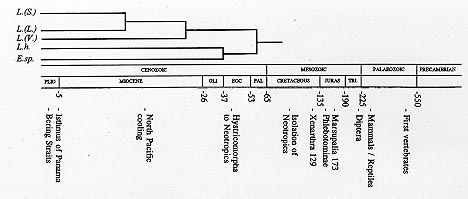
Vol. 93 (5), September/October 1998, pp. 657-662 Implications of a Neotropical Origin of the Genus Leishmania Harry Noyes Liverpool School of Tropical Medicine, Pembroke Place, Liverpool, L3 5QA, UK Received 15 June 1998; Accepted 30 July 1998
Code Number:OC98125
The hypothesis of a Neotropical origin of the Leishmania/Endotrypanum clade is reviewed. The position of the L. (Sauroleishmania) external to the subgenus L. (Leishmania) is not consistent with the Neotropical origin of the latter subgenus. It is suggested that this may be a consequence of a faster evolutionary rate in the L. (Sauroleishmania). The implications for the classsification of the phlebotomine sandflies of the hypothesis for a Neotropical origin of the Leishmania is also considered. The classification of Galati (1995) is proposed to be most consistent with the hypothesis of a Neotropical origin of the Leishmania, whilst classifications which place the New and Old World species in separate taxa are inconsistent with this hypothesis. Key words: Endotrypanum - Leishmania (Sauroleishmania) - Leishmania hertigi - Phytomonas - porcupines - sloths - Phlebotominae - biogeography FULL TEXT In recent years DNA sequence based phylogenies have transformed our understanding of the evolutionary relationships amongst a wide range of protozoa. The growing number of taxa examined and the increasing range of molecules used now makes it possible to consider how host parasite systems have coevolved with more confidence. The family Trypanosomatidae consists of nine genera of parasitic flagellated protozoa. Five of these genera are parasites of arthropods only and are transmitted contaminatively (Wallace 1979). Three genera have digenetic lifecycles in vertebrates and invertebrates and one genus the Phytomonas is parasitic in plants and insects. Phylogenies of the ribosomal RNA genes of members of the Trypanosomatidae indicate that vertebrate parasitism has arisen on at least two separate occasions within this family, once in the Trypanosoma and once in the Leishmania/Endotrypanum clade (Fernandes et al. 1993, Hollar & Maslov 1997, Lukes et al. 1997, Noyes 1998). The increasing number of taxa used in these phylogenies makes it possible to calibrate these phylogenies against specific events in the evolution of the hosts of these parasites and hence to consider how host parasite systems may have coevolved through time. The genus Trypanosoma is a cosmopolitan parasite of almost all classes of vertebrates and may have very ancient origins in the Palaeozoic. The Trypanosoma that are infective to humans may be of much more recent origin in the continents in which they are now found (Stevens et al. 1998). The genus Phytomonas is parasitic in flowering plants to which it is transmitted by a range of Hemiptera and Diptera. Since the first flowering plants appeared in the fossil record in the early Cretaceous (130 million years ago - MYA) and began to dominate the terrestrial flora during the second half of the Cretaceous it is possible that the genus Phytomonas made the transition from monogenetic parasites of plant feeding insects to digenetic parasites of plants and insects during the Cretaceous. The remaining two genera of digenetic parasites the Endotrypanum and the Leishmania are the two most closely related genera in the rRNA phylogenies of the Trypanosomatidae and their common ancestor may have made a transition to digenetic parasitism around the time of the mammalian radiation in the late Cretaceous or early Cenozoic (Fernandes et al. 1993). Leishmania and Endotrypanum are both transmitted by phlebotomine sandflies but the genus Endotrypanum only infects sloths in the Neotropics whilst the genus Leishmania infects at least nine orders of mammals and reptiles and is found in the tropics and subtropics worldwide. Recent phylogenies of the Leishmania/Endotrypanum clade have shown that the parasites which are endemic in the New World are closer to the root of this clade than the Old World parasites (Fig. 1). Consequently it has been proposed that this clade made the switch from monogenetic parasites of phlebotomine sandflies to digenetic parasites of sandflies and vertebrates in the Neotropics (Croan et al. 1997, Noyes et al. 1997). This switch is believed to have occurred at the time of the mammalian radiation at the end of the Cretaceous when the Neotropics were becoming isolated from the rest of the world (Fig. 2). At this time only four groups of mammals are known to have been established in the Neotropics: the monotremes, the marsupials, the notoungulates and the xenarthrans (Fig. 1). The monotremes and notoungulates became extinct in the Neotropics during the Cenozoic leaving the marsupials and xenarthrans as the only indigenous mammals to survive in the Neotropics to the present day (Patterson & Pascual 1972). Figure 1: geological periods plotted on a logarithmic scale showing the approximate dates of the appearance of mammal and insect groups important for the understanding of the evolution of the Endotrypanum/Leishmania clade. Numbers indicate millions of years before the present. Dates of divergence of mammal groups are from Kumar and Hedges (1998). A phylogeny of the Endotrypanum/ Leishmania clade is shown based on the phylogeny of RFLPs of the small subunit ribosomal RNA gene of Noyes et al. (1997). The phylogeny is calibrated against the divergence of the L. hertigi and Endotrypanum clades which is assumed to have occurred no earlier than the late Eocene at the time the Hystricomorph rodents first appeared in the Neotropical fossil records. L. (S.): L. (Sauroleishmania); L. (L.): L. (Leishmania); L. (V.): L. (Viannia); L. h.: L. hertigi; E. sp.: Endotrypanum sp. Figure 2: a map of the world in the late Cretaceous (65-100 million years ago - MYA) showing the isolation of the Neotropics from the Palaearctic. The Isthmus of Panama did not reconnect the two continents until 5 MYA in the Pliocene. The Beringia region which connected the Palaearctic to the Nearctic for most of the Cenozoic (0-65MYA) is shown. The shaded areas indicate shallow seas that covered large parts of the Palaearctic and Nearctic during the Mesozoic and Cenozoic (after Cox 1973). The Xenarthra consists of the sloths, the armadillos and the anteaters. The two genera of modern sloths Bradypus and Choloepus belong to two different families which diverged at the end of the Cretaceous over 70 MYA (Höss et al. 1996). The modern genera are the remnants of a much more diverse group of terrestrial, arboreal and even aquatic sloths that flourished during most of the Cenozoic (Patterson & Pascual 1972, Engelmann 1985, de Muizon & McDonald 1995). Since the two modern genera of sloths are hosts to both genera within the Leishmania/Endotrypanum clade it is possible that sloths were the first vertebrate hosts of the common ancestor of these parasites. Monogenetic trypanosomatid parasites of sandflies are not uncommon, although they are rarely described. These trypanosomatids may have become preadapted to development in the blood of vertebrates by frequent exposure to blood in the gut of their sandfly hosts. The relatively low body temperature of modern sloths, and presumably extinct sloths as well, may have facilitated the switch from sandfly to vertebrate hosts. Alternatively it has been suggested that since a number of genera of monogenetic and digenetic trypanosomatids can flourish in the anal scent glands of marsupials, that this organ provided a transitional stage in the acquisition of vertebrate parasitism and that consequently marsupials were the primary hosts of the Leishmania/Endotrypanum clade (Deane & Jansen 1988, Jansen et al. 1988). However the sandfly gut is an environment in which the parasite is more likely to be routinely exposed to vertebrate blood over many generations, than in the marsupial anal gland where the parasites have a lower chance of onward transmission. It may be difficult to resolve this question until more is known of the lifecycle and host specificity of the monoxenous trypanosomatids of sandflies. The phylogenies of the Leishmania/Endotrypanum clade show that the L. hertigi complex is more closely related to the genus Endotrypanum than to the Leishmania (Fig 1.) (Croan et al. 1997, Noyes et al. 1997). Since parasites of the L. hertigi complex have only been found in Neotropical porcupines (Rodentia: Hystri-chomorpha: Erethizontidae) it is possible that these parasites did not diverge from the Endotrypanum line until the hystricomorph rodents arrived in the New World during the Eocene. This provides a calibration point for dating the remaining nodes in the Endotrypanum/Leishmania tree (Fig.1). This suggests that the Endotrypanum and Leishmania genera diverged during the Palaeocene 53-65MYA, the Neotropical subgenus L. (Viannia) branched off during the early Miocene and the L. (Leishmania) and L. (Sauroleishmania) diverged during the second half of the Miocene. The tree used to estimate these dates was prepared from RFLPs of the small subunit rRNA gene (Noyes et al. 1997). Further trees including more taxa and data from more genes should increase the reliability of these estimates. Since the subgenus L. (Leishmania) is found in both the Old and New Worlds a member of this subgenus may have migrated to the Old World across the Bering straits region before this region became too cool for the sandfly vectors in the late Miocene (Wolfe 1994). The subgenus L. (Sauroleishmania) may then have diverged from the L. (Leishmania) in the Old World as a consequence of its adaption to reptiles. Although this hypothesis for the origin of the L. (Sauroleishma-nia) in the Old World requires the minimum number of migrations and extinctions it is not consistent with the phylogenies. If the L. (Sauroleishma-nia) had evolved in the Old World from L. (Leishmania) parasites that had migrated from the New World then the RNA and DNA polymerase phylogeny of Croan et al. (1997) would be expected to show the L. (Sauroleishmania) branching off between the L. (L.) mexicana complex which is restricted to the New World and the L. (L.) major, L. (L.) tropica and L. (L.) donovani complexes which are restricted to the Old World. Instead it shows that L. (Sauroleishmania) branched off before any of the L. (Leishmania) subgenus. It is possible that either the L. (Sauroleishmania) first evolved in the New World and then also migrated to the Old World independently of the L. (Leishmania) before becoming extinct in the New World, or that the common ancestor of both subgenera migrated to the Old World and that the ancestors of L. mexicana migrated back after the L. (Sauroleishmania) had diverged. However it seems more likely that the position of the L. (Sauroleish-mania) external to all the L. (Leishmania) in the RNA and DNA polymerase phylogeny is an artefact of a faster evolutionary rate in the L. (Sauroleishmania). The long length of the branch leading to the L. (Sauroleishmania) is suggestive of a faster evolutionary rate a possibility that was also indicated by a rate test (Croan et al. 1997). Faster evolving groups are known to be pulled towards the outgroup, a phenomenon known as long branch attraction, which could have generated the observed phylogeny (Felsenstein 1988). It is conceivable that the changes that were necessary for the L. (Sauroleishmania) to adapt from mammalian hosts to reptile hosts may have forced a temporarily accelerated rate of evolution. It may be possible to test this hypothesis more rigorously by the inclusion of additional taxa in this phylogeny. It is still not known whether the L. (Sauroleish-mania) are transmitted by the bite of the sandfly or by contamination when the reptile eats the sandfly (Telford 1995). Since it now appears that the L. (Sauroleishmania) have evolved from parasites that are transmitted by the bite of the fly this is perhaps the most likely method for transmission of L. (Sauroleishmania) to reptiles as well. Most Leishmania parasites are more restricted in their range of sandfly vectors than in their range of mammalian hosts, implying a much closer coevolutionary relationship with the sandfly than the mammal. However the proposed Neotropical origin of the Leishmania is not consistent with the existing classification of the sandflies. Nevertheless relationships between the subgenera and species complexes of sandflies are still controversial and the existing nomenclature may not reflect the true relationships within this group (Lane 1993). If the hypothesis for a Neotropical origin is correct then it will be possible to make some predictions for the classification of the sandflies that could be tested by molecular methods. In the Old World mammalian Leishmania are transmitted by sandflies of the genus Phlebotomus and lizard parasites are transmitted by sandflies of the genus Sergentomyia. In the New World Leishmania and Endotrypanum are transmitted by sandflies of the genus Lutzomyia (Fig. 3). If Leishmania migrated across the Bering region during the Miocene there must have been a resident population of sandfly vectors throughout this area which may have left descendants in both the Old and New Worlds. Consequently the modern sandfly vectors in both the Old and New World may be more closely related to each other than they are to sympatric non-vector genera. Figure 3: the modern distribution of sandfly vector genera, after Lewis (1974) and Leishmania. The proposed route by which Leishmania migrated from the Neotropics to the Old World is indicated by an arrow. The distribution of sandflies is limited by summer temperatures which must remain above 20°C for 50 days a year (Lewis 1982). Areas with a mean temperature of less than 20°C in the hottest month (June for the Northern hemisphere and January for the Southern Hemisphere) are shaded. These areas are not normally suitable inhabited by Phlebotominae. In the Northern Hemisphere the 20°C isotherm is at about 45°N, for the Beringia region to be suitable One recent phylogeny of the sandflies does suggest that this is the case. Galati (1995) places the Old World genus Sergentomyia in a new subtribe, the Sergentomyiina, with some reptile biting species that are at present in the New World genus Lutzomyia. In this classification the Sergentomyiina is clustered in a group of New World subtribes which suggests that the Sergentomyiina may also have evolved in the New World and that the modern Sergentomyia are descendants of Sergentomyiina that migrated from the New World to the Old World. This implies that sandflies could have crossed through Beringia at some time. The Sergentomyiina are primarily reptile biters, but members of the Lutzomyia vexator series, which Galati places within this subtribe, have been implicated as the vectors of L. mexicana in Texas (Kerr et al. 1995). Consequently it is possible that ancestors of the widespread Lu. vexator series or of a closely related group may have carried the mammalian Leishmania to the Old World. No molecular phylogenies of the Phlebotominae have been published, but they will provide a valuable test of the hypothesis for a Neotropical origin of the Leishmania. REFERENCES Cox CB 1974. Vertebrate palaeodistributional patterns and continental drift. J Biogeog 1: 75-94. Croan DG, Morrison DA, Ellis JT 1997. Evolution of the genus Leishmania revealed by comparison of DNA and RNA polymerase gene sequences. Mol Biochem Parasitol 89: 149-159. de Muizon, C, McDonald, HG 1995. An aquatic sloth from the Pliocene of Peru. Nature 375: 224-227 Deane MP, Jansen AM 1988. From a mono to a digenetic life-cycle: how was the jump for flagellates of the family trypanosomatidae. Mem Inst Oswaldo Cruz 83: 273-275. Engelmann GF 1985. The phylogeny of the Xenarthra, p. 51-64. In GG Montgomery, The Evolution and Ecology of Armadillos, Sloths and Vermilinguas, Smithsonian Institution Press, Washington. Felsenstein J 1988. Phylogenies from molecular sequences: inference and reliability. Annu Rev Genet 22: 521-565. Fernandes AP, Nelson K, Beverley SM 1993. Evolution of nuclear ribosomal RNAs in kinetoplastid protozoa - perspectives on the age and origins of parasitism. Proc Natl Acad Sci USA 90: 11608-11612. Galati BEA 1995. Phylogenetic systematics of Phlebotominae (Diptera, Psychodidae) with emphasis on American groups. Boletín de la Direccion de Malariologia y Saneamiento Ambiental 35 (Suppl. 1): 133-142. Höss M, Dilling A, Currant A, Paabo S 1996. Molecular phylogeny of the extinct ground sloth Mylodon darwinii. Proc Natl Acad Sci USA 93: 181-185. Hollar L, Maslov DA 1997. A phylogenetic view on the genus Phytomonas. Mol Biochem Parasitol 89: 295-299 Jansen AM, Carreira JC, Deane MP 1988. Infection of a mammal by monogenetic insect trypanosomatids (Kinetoplastida, Trypanosomatidae). Mem Inst Oswaldo Cruz 83: 271-272. Kerr SF, McHugh CP, Dronen Jr NO 1995. Leishmaniasis in Texas: prevalence and seasonal transmission of Leishmania mexicana in Neotoma micropus. Am J Trop Med Hyg 53: 73-77. Kumar S, Hedges SB 1998. A molecular timescale for vertebrate evolution. Nature 392: 917-920 Lane RP 1993. Sandflies (Phlebotominae), p. 78-119. In RP Lane & RW Crosskey (eds), Medical Insects and Arachnids, Chapman & Hall, London. Lewis DJ 1974. The biology of Phlebotomidae in relation to leishmaniasis. Ann Rev Entomol 19: 363-384. Lewis DJ 1982. A taxonomic review of the genus Phlebotomus (Diptera: Psychodidae). Bull Br Mus Nat Hist (Ent) 45: 121-209. Lukes J, Jirku M, Dolezel D, Kral'ová I, Hollar L, Maslov D 1997. Analysis of ribosomal RNA genes suggests that trypanosomes are monophyletic. J Mol Evol 44: 521-527. Noyes HA 1998. Can Trypanosoma trees be trusted? Parasitol Today 14: 49-50. Noyes HA, Arana BA, Chance ML, Maingon R 1997. The Leishmania hertigi (Kinetoplastida; Trypanosomatidae) complex and the lizard Leishmania: their classification and evidence for a neotropical origin of the Leishmania-Endotrypanum clade. J Eukaryot Microbiol 44: 511-517. Patterson B, Pascual R 1972. The fossil mammal fauna of South America, p. 247-310. In A Keast F Erk & B Glass (eds), Evolution, Mammals and Southern Continents, State University of New York Press, Albany. Stevens JR, Noyes HA, Dover GA, Gibson WC 1998. The ancient and divergent origins of the human pathogenic trypanosomes, Trypanosoma brucei and T. cruzi. Parasitology, in press. Telford Jr SR 1995. The kinetoplastid hemoflagellates of reptiles, p. 161-223. In JP Kreier, Parasitic Protozoa, 2nd ed., Vol. 10, Academic Press, San Diego. Wallace FG 1979. Biology of the Kinetoplastida of arthropods, p. 213-240. In WHR Lumsden & DA Evans (eds), Biology of the Kinetoplastida, Vol. 2, Academic Press, London. Wolfe JA 1994. An analysis of Neogene climates in Beringia. Palaeogeogr Palaeoclimatol Palaeoecol 108: 207-216. Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98125c.jpg] [oc98125b.jpg] [oc98125a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}