 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
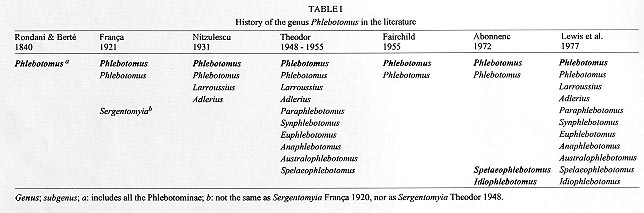
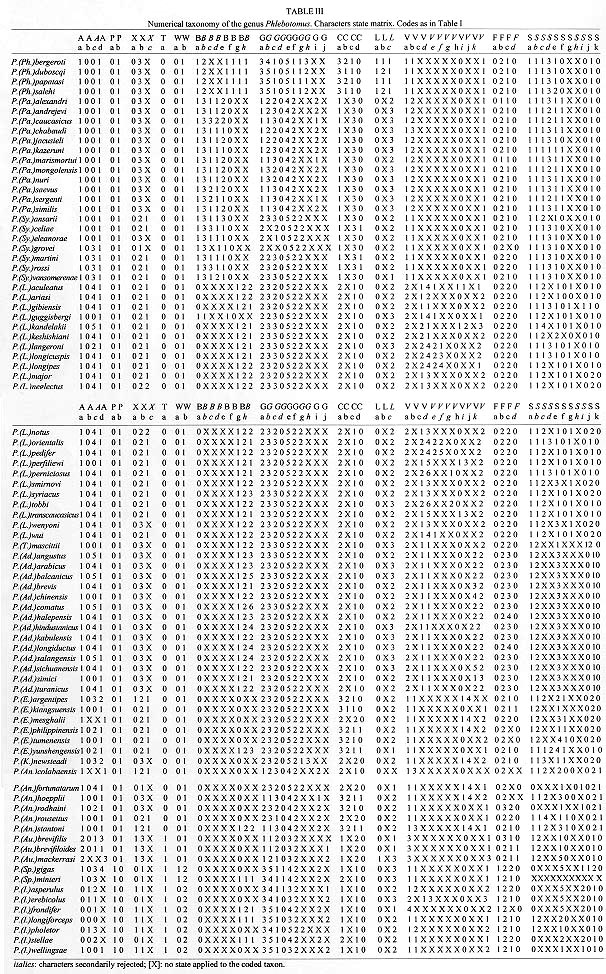
Vol. 93(6): 773-785 Numerical Taxonomy of Old World Phlebotominae (Diptera: Psychodidae). 1. Considerations of Morphological Characters in the Genus Phlebotomus Rondani & Berté 1840 Philippe Rispail/+, Nicole Léger* Laboratoire d'Ecologie Médicale et Pathologie Parasitaire (Pr J-P Dedet),
Faculté de Médecine, 163 rue Auguste-Broussonet, 34090 Montpellier, France
Received 26 March 1998; Accepted 29 July 1998
Code Number:OC98210
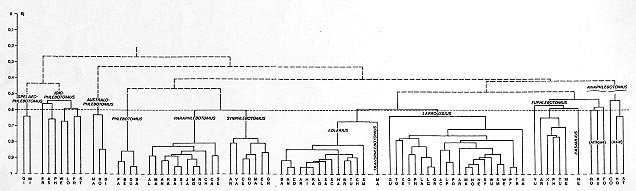
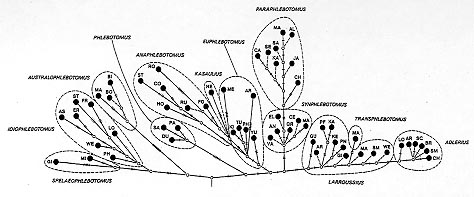
Numerical analyses (correspondence analysis, ascending hierarchical classification, cladistic approach) were applied to the morphological characters of the adults of the genus Phlebotomus Rondani & Berté 1840. They confirm the reliability of the classic classifications, and also redefine the taxonomic and phylogenetic position of certain taxa. Thus, Spelaeophlebotomus Theodor 1948, Idiophlebotomus Quate & Fairchild 1961 and Australophlebotomus Theodor 1948 deserve generic rank. Among the vectors of leishmaniasis, the subgenus Phlebotomus Rondani & Berté 1840 is probably ancient. The results attribute an intermediate taxonomic and phylogenetic position to the taxa Euphlebotomus Theodor 1948 and Anaphlebotomus Theodor 1948, and reveal the probable artificial nature of the latter. The comparatively large numbers of species of subgenera Paraphlebotomus Theodor 1948, Synphlebotomus Theodor 1948 and, above all, Larroussius Nitzulescu 1931 and Adlerius Nitzulescu 1931, suggest that they are relatively recent. The development of adult morphological characters, the validity of their use in taxonomy and proposals for further studies are discussed. Key words: Old World sand flies - Phlebotomus - Psychodidae Phlebotominae - numerical taxonomy Taxonomy of the genus Flebotomus, created by Rondani and Berté (Rondani 1840) and emended to Phlebotomus by Loew (1845), has relied on a small number of adult characters considered separately but on a priori grounds given discriminant values (monothetic taxonomy). During more a century, the genus has been subdivided into an increasing number of subgenus ( Table I). In 1982, Lewis subdivided Phlebotomus into 11 subgenera to which Abonnencius UbedaOntiveros et al. 1982 and Transphlebotomus Artemiev & Neronov 1984 were later added. The former was synonymised with Anaphlebotomus Theodor 1948 by Lane and Alexander (1988). Some authors gave generic rank to Spelaeophlebotomus Theodor 1948, Idiophlebotomus Quate & Fairchild 1961 and Australophlebotomus Theodor 1948. There is some uncertainty as to the appropriate rank for these taxa and their phylogeny, and the evolutionary direction of the characters used by systematists. Numerical polythetic taxonomy simultaneously takes into account numerous characters without according any discriminant value, using advancements in information technology. Phenetic analysis expresses the intergroup relationships by means of factorial graphs and dendrograms. Cladistic analysis ends in the construction of cladograms, permitting the erection of filiation hypotheses and judgement of the direction of development of the characters. MATERIALS AND METHODS Selection of taxa - For the numerical analysis, specific taxa were chosen as the operational taxonomic units (OTU) (Sokal & Sneath 1963). An examination of the available specimens permitted us to select discriminant characters. Lewis (1982) considered that the genus Phlebotomus Rondani & Berté 1840 was composed of about 110 specific or infraspecific taxa, unevenly grouped into 12 subgenera. Four arguments led to the elimination of certain taxa from the numerical analysis: validity does not seem to be totally established on solid bases (morphological as well as systematic); subspecific position is probable; inevitable constraints on the coding of the characters in closely related taxa; finally, and above all, a crippling imbalance in the calculation tables caused by missing data (incomplete description, species of which only one sex is known). Because of the latter the following taxa were eliminated: P. (Idiophlebotomus) sejunctus Quate 1965, P. (Id.) teshi Lewis 1978 and P. (Id.) tubifer Lewis & Lane 1976; P. (Australophlebotomus) acuminatus Lewis & Dyce 1982, P. (Au.) buccinator Fairchild 1952, P. (Au.) papuensis Fairchild 1952, P. (Au.) pexopharynx Fairchild 1952 and P. (Au.) trifilis Quate & Quate 1967; P. (Synphlebotomus) katangensis Bequaert & Walravens 1930 and P. (Sy.) taylori Davidson 1982; P. (Larroussius) betisi Lewis & Wharton 1963, P. (La.) chadlii Rioux, Juminer & Gibily 1966, P. (La.) fantalensis Lewis, Minter & Ashford 1974 and P. (La.) mariae Rioux, Croset, Léger & BaillyChoumara 1974; P. (Euphlebotomus) autumnalis Artemiev 1980 and P. (Eu.) caudatus Artemiev 1979; P. (Anaphlebotomus) somaliensis Abonnenc, Adam & BaillyChoumara 1959. Since our work begun, the female of P.(L.) fantalensis Lewis, Minter & Ashford 1974 has been described. Three new species of the subgenus Larroussius have been named: P.(L.) lengi Zhang, He & Ward 1994, P.(L.) ashfordi Gebre-Michael & Lane 1996 and P.(L.) mireillae Killick-Kendrick et al. 1997. More, P.(L.) elgonensis Ngoka, Madel & Mutinga 1975 has been taken reinstated (Killick-Kendrick et al. 1993). Eventually 85 OTU's were retained. A preliminary cladistic analysis carried out jointly with phenetic analysis resulted in the establishment of the evolutionary direction and assessment of the validity of the characters. Twentythree characters had to be eliminated, making the specific coding of each of the 85 taxa impossible, therefore 26 OTU's had to be excluded. Choice of characters and definition of states -The arguments of choice were: availability for all the taxa, absence of redundancy, clear definition of the states of characters and the stability of these states at a taxonomic level. Quantitative characters were, for the most part, excluded because of their variability. The description of the 85 taxa required the use of 63 characters and 176 character states (Table II). Numerical analysis - Following the tabulation of character states for each OTU (Table III), analyses were carried out using an Olivetti M380 XP1 computer. Phenetic analysis - Programmes were developed from those of the algorithm Analyse de Données of the software Biomeco 2.0 package (Groupe Biométrie CEPE/CNRS, Montpellier). Similarity tables of distance indices (Jaccard 1908) were transformed into multidimensional scatter diagrams, reduced to two dimensions by factor analysis, or into dendrograms, by ascending hierarchical classification. In the latter, cluster analysis used intermediate linkage, and, in the absence of additional information, linkages at less than 50% similarity were considered as random. Cladistic analysis - The MIX algorithm (Wagner parsimony), extracted from the PHYLIP programme, distributed by Felsenstein (1978, 1985), was used. This allows differentiation between occasional missing values and indeterminations, expressing characters of which no state applies to the coded taxon. Bias introduced by concentration of characters relating to a single organ is compensated by ascribing reduced weight to dependent characters. In the absence of sufficiently well known fossils to determine plesiomorphy, the supposed evolutionary direction of the greatest number of these characters, required to root the phylogenetic tree, was supported from studies of other Psychodidae (Hennig 1972). The construction of Wagner diagrams (Wagner 1961) used Manhattan distance (Legendre & Legendre 1979), and took account simultaneously of the character states and the distances between OTU's. RESULTS Phenetic analysis The first axis of the correspondence analysis separates the species of the taxa Idiophlebotomus, Spelaeophlebotomus and Australophlebotomus from those of the others groups. The most important characters in this axis are the antennal formula in the female, the palpal formula and the morphology of the sensillae, the presence or absence of mesanepisternal setae, the shape of the wing, the structure and number of spines on the style, the presence or absence of intraabdominal rods, and the morphology of the spermathecae. The second axis separates Paraphlebotomus and Synphlebo-tomus from Idiophlebotomus and Spelaeophlebo-tomus. The important characters here are the male genital structure (basal lobe, style, paramere, aedeagus) and the presence or absence of segmentation on the spermathecae. As with the correspondence analysis, ascending hierarchical classification (Fig. 1) confirms generally accepted subgenera. While the subgeneric level overall is approximately 60% of similarity, the species of the subgenera Phlebotomus, Paraphlebotomus, Synphlebotomus, Adlerius and Larroussius join at a high level of similarity (7080%). The species of the other taxa are further one from the other. Paraphlebotomus and Synphlebotomus come together early, then are joined by Phlebotomus. P. (Transphlebotomus) mascittii attaches to Adlerius before the major union with Larroussius. The numerous distinctive characters of P. (Euphle-botomus) yunshengensis integrate P. (Kasaulius) newsteadi into the block of Euphlebotomus. The three African species of Anaphlebotomus join to Euphlebotomus, while the three Asiatic species are completely separate. The taxa Spelaeophleboto-mus, Idiophlebotomus and Australophlebotomus separate very late, at a level of similarity without any significance. Fig. 1: 85 species of the genus Phlebotomus. Ascending hierarchical classification using 63 characters and 176 states. Dendrogram constructed using Jaccard's similarity index and clustering by median linkage. Spelaeophlebotomus. GI: P. gigas MI: P. minteri. Idiophlebotomus. AS: P. asperulus ER: P. erebicolus FR: P. frondifer LO: P. longiforceps PH: P. pholetor ST: P. stellae WE: P. wellingsae. Australophlebotomus. BI: P. brevifilis BO: P. brevifiloides MA: P. mackerrasi. Phlebotomus. BE: P. bergeroti DU: P. duboscqi PA: P. papatasi SA: P. salehi. Paraphlebotomus. AL: P. alexandri AN: P. andrejevi CA: P. caucasicus CH: P. chabaudi JA: P. jacusieli KA: P. kazeruni MA: P. marismortui MO: P. mongolensis NU: P. nuri SA: P. saevus SE: P. sergenti SI: P. similis. Synphlebotomus. AN: P. ansarii CE: P. celiae EL: P. eleanorae GR: P. grovei MA: P. martini RO: P. rossi VA: P. vansomerenae. Adlerius. AN: P. angustus AR: P. arabicus BA: P. balcanicus BR: P. brevis CH: P. chinensis CO: P. comatus HA: P. halepensis HI: P. hindustanicus KA: P. kabulensis LO: P. longiductus SA: P. salangensis SC: P. sichuanensis SM: P. simici TU: P. turanicus. Transphlebotomus. MA: P. mascittii. Larroussius. AC: P. aculeatus AR: P. ariasi GI: P. gibiensis GU: P. guggisbergi KA: P. kandelakii KE: P. keshishiani LA: P. langeroni LC: P. longicuspis LP: P. longipes MA: P. major NE: P. neglectus NO: P. notus OR: P. orientalis PD: P. pedifer PF: P. perfiliewi PN: P. perniciosus SM: P. smirnovi SY: P. syriacus TO: P. tobbi TR: P. transcaucasicus WE: P. wenyoni WU: P. wui. Euphlebotomus. AR: P. argentipes KI: P. kiangsuensis ME: P. mesghalii PH: P. philippinensis TU: P. tumenensis YU: P. yunshengensis. Kasaulius. NE: P. newsteadi. Anaphlebotomus. CO: P. colabaensis FO: P. fortunatarum HO: P. hoepplii RO: P. rodhaini RU: P. rousettus ST: P. stantoni. Cladistic analysis - Phylogenetic hypotheses and development of the characters First phylogenetic hypothesis - On the most parsimonious Wagner's tree achieved (Fig. 2), the species of the taxa Spelaeophlebotomus and Idiophlebotomus are the first to form a group. The species of the subgenera Australophlebotomus and Phlebotomus quickly diverge from an early common branch. The next branch contains the species of the subgenus Anaphlebotomus. The three African species diverge very quickly from the Asiatic branch. The branch Kasaulius comes from the Euphlebotomus branch. The first node in the branch appearing afterwards would be a hypothetical common ancestor of Synphlebotomus and Paraphle-botomus. The terminal branches illustrate the abundant specific diversification within the subgenera Larroussius, Transphlebotomus and Adlerius, which are probably the most developed. Fig. 2: 85 species of the genus Phlebotomus. Cladistic analysis using 63 characters and 176 states. Most parsimonious cladogram (317 steps). Symbols are the same as in Fig. 1. Bars indicate the number of steps. Evolution of the characters - One of the major interests of cladistic analysis is the
consideration of each character individually: definition of its states, probable direction of
development and the validity of its use. To investigate this, each one of the 63 characters used
was followed from the root of the tree to its last branches. The principal conclusions from this
study are: Antennae - Accepting the presence of two ascoids on antennal segments III
to XVI in the female, as plesiomorphic by analogy with the other Psychodidae,
Idiophlebotomus appears to be an ancestral group, Australophlebotomus
seems be very well developed and Spelaeophlebotomus is intermediate. On the other
hand, in the male, only three species of the genus Idiophlebotomus show the derivative
state "absence of two ascoids on the third segment", a condition which disagrees with that of
the antennal formula in the female, diminishing the cladistic interest of this character. The
papillary formula of the antennae 1/IIIIVV occurs only in the four species of the branch
Phlebotomus and in P. (Australophlebotomus) brevifiloides.
However, the lack of data for the majority of the species of the taxa
Spelaeophlebotomus and Idiophlebotomus prevents us from confirming that
this state is really ancestral. Palps - The plesiomorphy of the absence of spatulate
sensillae found only in Spelaeophlebotomus and Idiophlebotomus has not been
formally confirmed, but is compatible with the structure of the phylogenetic tree. Palpal
segments 3 longer than the others are seen in the group Spelaeophlebotomus
Idiophlebotomus which is probably ancestral. Thus this state seems to be
plesiomorphic. Cibarium - The cibarium of the female is armed with teeth only in
Australophleboto-mus and the majority of Idiophlebotomus. The presence of a
well developed armature in the neighbouring genera Sergentomyia and
Lutzomyia leads us to believe that the analysis of this character should be done at a
family level. Pharynx - The evolution of the pharyngeal armature seems to be in the
direction of the development of structures from simple ridges towards large teeth with forward
expansion. As the most well developed state is always found at the end of the branch, the
development of the pharyngeal armature is probably an adaptative character as, perhaps, is the
cibarial armature. Thorax - Following the examples of the American taxa
Hertigia and Warileya, alone within the genus Phlebotomus, the
species of the taxa Spelaeophlebotomus and Idiophlebotomus do not have an
anteroinferior cluster of mesanepisternal bristles or scales. If one considers these groups as
more primitive than the others, the hypothesis of a development towards the disappearance of
this cluster must be submitted to further investigation. In fact the discussion essentially
concerns whether or not these two taxa belong in the genus Phlebotomus.
Wings - In Spelaeophlebotomus the fourth vein (M1+M2) branches at the
level of the radiomedial (rm). In all the other taxa, this branch is more or less distant from rm.
Relatively short and wide wings are characteristic of the primitive groups, and the hypothesis
of evolutionary lengthening of "gamma" seems to be verified. A ratio "width of wing/gamma"
equal to or greater than 4 is apparently an ancestral character present in the taxa
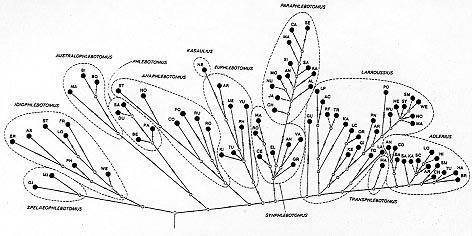
Spelaeophlebotomus and Idiophlebotomus. Genital pump and filaments - Exceptionally small or large genital pumps are apomorphic states. Development was presumably through the progressive increase in the ratio length of genital filaments / length of the pump. From the ancestral state "less than 3", it increases to "35" in Larroussius and Transphlebotomus, to "6.59" then "greater than 9.5" in Adlerius. Lateral lobes - The absence of spines on the lateral lobes is confirmed as a plesiomorphic state. The apomorphic state only concerns the apical spatulate spines of Phlebotomus. The number of spines increases from two in P. bergeroti and P. papatasi to more than three in P. duboscqi and P. salehi. Spermathecae - Cylindrical spermathecae seem to be ancestral. The presence of ornamentation on the reservoir wall of the spermathecae is exclusive to the developed groups: only the taxa Spelaeophlebotomus, Idiophleboto-mus and Anaphlebotomus contain the species with smooth spermathecae. In these groups, the evolution from smooth, poorly defined spermathecae towards spermathecae with a well defined capsule cannot be confirmed. In the species with ornamented spermathecae, clear segmentation has progressively diminished. The overall development would, therefore, be from the smooth spermathecae to the clearly segmented spermathecae and then to the pleated or ridged spermathecae. The separate opening of the spermathecae ducts is a plesiomorphic character. Common ducts occur at the end of the branches: in P. (Sp.) gigas, Anaphlebotomus, Kasaulius and Euphlebotomus (except P. yunshengensis), P. (T.) mascittii and the major group within the subgenus Larroussius. It must be noted that the demonstration of this state requires delicate dissection and it is not possible to be sure that this has been carried out with all the species. Moreover, within the genus Phlebotomus, exceptionally long ducts are a synapomorphy of the subgenus Anaphlebotomus. Cylindrical spermathecae, without ornamentation, and with ducts of uniform diameter opening separately are, therefore, very likely to be close to the ancestral spermathecal structure of the group. Second hypothesis - This analysis of the characters has led to the abolition (or in one case the regrouping) of certain characters. This reduction in the number of 63 characters to 40, subdivised into 102 states, brings the number of individual species considered to 59. Despite this loss of data at the specific level, the overall structure of the new phylogenetic tree is not seriously altered (Fig. 3). Therefore the abolished characters have little influence on the first phylogenetic hypothesis. Some noticeable differences are, however, to be found. The taxa Spelaeophlebotomus, Idiophlebotomus and Australophlebotomus constitute one of the two initial branches of the tree. All of the known vector species of leishmaniasis are situated on the other branch. The branch Phlebotomus arises very early. Anaphlebotomus emerges from Euphlebotomus in two distinct branches. The subgenera which are probably the most developed, Synphlebotomus and Paraphlebotomus, then Larroussius, Transphlebo-tomus and Adlerius, divide up in the same way as on the previous tree. Fig. 3: 59 species of the genus Phlebotomus. Cladistic analysis using 40 characters and 102 states. Most parsimonious cladogram (163 steps). Symbols are the same as in Fig. 1. Bars indicate the number of steps. Elimination of certain characters prevents discrimination between certain taxa: P.(Ph.) bergeroti and P.(Ph.) papatasi; P.(Pa.) andrejevi, P.(Pa.) mongolensis, P.(Pa.) nuri, P.(Pa.) similis and P.(Pa.) saevus; P.(Sy.) rossi and P.(Sy.) martini; P.(L.) aculeatus, P.(L.) transcaucasicus and P.(L.) perfiliewi; P.(L.) langeroni, P.(L.) longicuspis, P.(L.) longipes, P.(L.) orientalis, P.(L.) pedifer, P.(L.) tobbi and P.(L.) perniciosus; P.(L.) neglectus, P.(L.) notus, P.(L.) syriacus, P.(L.) wui and P.(L.) major; P.(Ad.) angustus, P.(Ad.) balcanicus, P.(Ad.) comatus, P.(Ad.) hindustanicus, P.(Ad.) kabulensis, P.(Ad.) salangensis, P.(Ad.) turanicus and P.(Ad.) arabicus; P.(Ad.) halepensis and P.(Ad.) longiductus. DISCUSSION Characters used and methodology - In future cladistic analysis, greater selectivity of characters is required, although the problem of separation of the closely related species will then become acute. Because of the taxonomic and phylogenetic importance of each group of characters, this choice would have to be reasoned and very prudent. Despite the technical balancing carried out, the characters probably do not carry equal weighting in the analysis. Thus, the probably adaptative characters should be accorded lesser evolutionary weighting. On the basis of their supposed development in related groups or at the level of the family, certain character states were considered a priori as ancestral. Some of these "postulats" were clearly invalidated by the cladistic analysis and the study of the characters within the genus. In order to root the phyletic tree, it is therefore necessary to limit the plesiomorphies at the beginning of the cladistic process, to those firmly established at the level of the family. In fact a study limited to the species of the genus Phlebotomus can only give a very partial insight into the development of certain characters in the Phlebotominae or in the Psychodidae as a whole. It is, therefore, essential to extend this study to other genera. Elsewhere, without hypothesizing plesiomorphies, the rooting of the tree by using of an outgroup such as Sergentomyia or another closely related Psychodid should be tried. Taxonomy and phylogeny - The methods of numerical analysis confirm the soundness of classical classifications, while refining the taxonomic levels. Spelaeophlebotomus and Idiophlebotomus - Whilst excluded from the genus Phlebotomus by some authors (Abonnenc 1972, Artemiev & Neronov 1984), these two taxa are retained as subgenera by others (Lewis et al. 1977, Lewis 1982), in the interests of stability. The phenetic classifications, factorial analysis or ascending hierarchical classifications, show a large distance between these two groups and the other subgenera. Also, the phylogenetic hypotheses show them to emerge precociously on a separate branch. The absence of spatulate sensillae on the palps, a ratio width of wing/gamma greater or equal to 4, and the long cylindrical styles are ancestral characters common to the species of these two taxa. They all possess three characters for which the developmental direction remains to be determined: long third palpal segments, absence of mesanepisternal setae, and presence of intraabdominal rods. Moreover Spelaeophlebotomus has an unarmed cibarium and a short gamma, probably plesiomorphic states. Furthermore, is the presence of only four spines on the styles, compensated for by a non deciduous bristle, really a developed state? Most of the species of the group Idiophlebotomus have a cibarial armature. The coxites of some species also have a group of bristles the subapical position of which is probably ancestral. Their respective position on the graphs, dendrograms or cladograms, as well as the detailed study of their characters, lead us to believe that Spelaeophlebotomus and Idiophle-botomus merit generic rank. Australophlebotomus - As with the last two, this subgenus clearly separates from the rest of the genus Phlebotomus. Phenetic analysis always positions it at a distance from the genus cluster, or on a isolated branch of the dendrogram. Initially linked to the subgenus Phlebotomus, it joins Spelaeo-phlebotomus and Idiophlebotomus on the same branch of the cladogram, after the elimination of certain characters. In the possession of long palpal segments 5 and some mesanepisternal bristles, Australophlebotomus is distinct from Spelaeophlebotomus and Idiophlebotomus. Furthermore, they have a number of characters in a developed state: female antennal formula, spatulate palpal sensillae, short ovoid styles with three spines, etc. Their phenetic distance from the other subgenera and the distinctive development of a number of the characters make Australophleboto-mus a clearly separate group which probably merits generic rank. Phlebotomus - This taxon (only four species) is placed amongst the more primitive subgenera. It is probably linked to Australophlebotomus. A certain number of characters shared with the known fossil species (morphology of the styles, grouping of the spines, etc.) support this hypothesis. The short spines on the styles and the spines on the lateral lobes are distinctive and probably apomorphic characters. Two other apomorphic characters, the small basal lobes and subapical group of bristles on the coxites, have a questionable taxonomic value, as their homology with those of more advanced groups requires further study. Euphlebotomus and Kasaulius - The interspecific distances within this group are comparatively large. As several distinctive characters of P. (K.) newsteadi (shape of the halteres, length of the legs, etc.) have not been taken into account, it is impossible totally to isolate this species from Euphlebotomus and, therefore, formally justify the creation of the subgenus Kasaulius Lewis 1982. The position of the two subgenera on the various cladograms suggests they are relatively ancient. Anaphlebotomus - Specific characters clearly split Anaphlebotomus into two groups, one Asian, the other African. The latter includes P. fortunatarum, which is native in the Canary Islands. In fact, the phenetic analysis reveals the probable artificial character of the subgenus. Cladistic analysis indicates that this is relatively ancient subgenus. Despite its lack of unity, it still seems to be the central region of the evolution of the genus. The relatively developed level of the group is, however, emphasised by the morphology of the styles (especially in the Asiatic species), by the presence of four spines (except in P. fortunatarum), and also by the existence of exceptionally long spermathecal ducts which join to form a common duct. Paraphlebotomus and Synphlebotomus - The phenetic analysis places these closely related subgenera near the subgenus Phlebotomus. On the other hand, the cladistic analysis situates them among the more highly evolved taxa, with recent speciation. The presence of long basal lobes with long bristles and the presence of spatulate tips to the parameres are developed characters common to them both. But the subgenus Paraphlebotomus seems to have evolved separately, developing generally short ovoid styles, with only four spines. Larroussius, Transphlebotomus and Adlerius - The phenetic proximity of these three subgenera is well established and the proliferation of the species within them is probably recent. While it is difficult to confirm the validity of the subgenus Transphlebotomus Artemiev & Neronov 1984, the taxa P. mascittii and P. canaaniticus, probably subspecies, seem closer to Adlerius than Larroussius. Their separation appears to be later than that of Larroussius, just before the appearance of Adlerius. A median group of bristles on the coxites and the long digitiform aedeagus, are developed characters common to all forms in this group. However, the particular case of P. (L.) guggisbergi must be noted; it has no group of coxal bristles, but, on each coxite, in the basal position, has a massive structure which is not very prominent, but is covered with bristles. Evolution in Larroussius included the development of a long digitiform extension of the spermathecal "neck". In the major of Larroussius group, as well as in Transphleboto-mus, the spermathecal ducts join to form a common duct. The ratio length of the male genital filaments/length of the genital pump ranges from 3 to 5 in Larroussius and Transphlebotomus, to at least 6.5 in Adlerius. The subterminal tubercle on the penis tips appears to be a recent acquisition in Adlerius. Outlook - Cladistic study should make use of characters not normally used in identification, including internal morphological features as well as larval and pupal characters. After careful selection of the characters, the study should be extended to other genera of the Phlebotominae. A similar process should be applied at the supraspecific level to Old World sand flies, secondarly to Psychodidae as a whole. This would perhaps enable the determination of a universally acceptable taxonomic status of sand flies: Phlebotomidae or Phlebotominae? Comparison of our results with the phenetic and cladistic analysis of biochemical and chromosomal data will make a major contribution to the taxonomy and phylogeny of the Phle-botominae. Eventual synthesis with ecological and behavioural data will allow a biogeographical and bioclimatic study to proceed on a firm basis. New light may be shed on populations and the evolution of characters. A combination of all this information, with parallel studies on Leishmania spp. and their reservoir hosts, will perhaps contribute to a fuller understanding of the structure of leishmaniasis foci. ACKNOWLEDGMENTS To Prof. F Vaillant (Faculté des Sciences, Grenoble), Prof. DM Jarry, Dr DT Jarry, Dr G Lanotte and Mr E Serres (Faculté de Médecine, Montpellier), Prof. RW Ashford (Liverpool School of Tropical Medicine), Dr L Matile and Dr JP Hugot (Muséum National d'Histoire Naturelle, Paris) for their valuable advice. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98210f.jpg] [oc98210a.jpg] [oc98210d.jpg] [oc98210b.jpg] [oc98210c.jpg] [oc98210e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}