 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 93(6): 795-798 RESEARCH NOTE A New Enzymatic Variant of Leishmania (Leishmania) forattinii Isolated from Proechimys iheringi (Rodentia, Echimydae) in Espírirto Santo, Brazil Aloisio Falqueto, Elisa Cupolillo*, Gerzia M Carvalho Machado*, Luiz Eduardo de Carvalho-Paes*, Gabriel Grimaldi Jr*/+ Departmento de Medicina Social, Centro Biomédico, UFES, Av. Marechal
Campos 1468, 29040-090 Vitória, ES, Brasil Received 29 April 1998; Accepted 28 July 1998
Code Number:OC98212
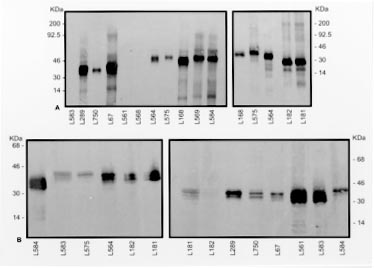
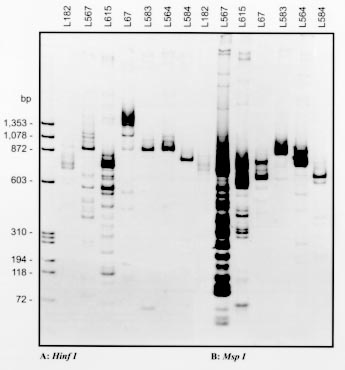
Key words: Leishmania - Leishmania (Leishmania) forattinii - taxonomy - monoclonal antibodies - isoenzyme electrophoresis - DNA analysis RESEARCH NOTE Taxonomic studies of leishmanial isolates from the New World indicate tremendous diversity within this genus (E Cupolillo et al. 1994 Am J Trop Med Hyg 50: 296-311). A number of new Leishmania species from sylvan areas of the Neotropics have been described recently. Some of those parasites are associated with disease in humans; others appear to be restricted to lower orders of mammals, such as rodents and edentates (G Grimaldi et al. 1989 Am J Trop Med Hyg 41: 687-725). However, some of the latter parasites may yet be shown capable of causing human disease, particularly in persons with altered cellular immune responses. During a survey of potential reservoir hosts of Leishmania done in the State of Espírito Santo, Brazil (1982-1985), three leishmanial parasites (designated with stock codes MPRO/BR/82/RV203, MPRO/BR/82/RV228, and MPRO/BR/85/RV260) were isolated from skin lesion samples taken from spiny-rats, Proechimys iheringi (Rodentia, Echimydae), captured in a secondary forest located in the municipality of Viana (A Falqueto et al. 1985 Mem Inst Oswaldo Cruz 80: 497). In studies using an indirect radioimmune binding assay (RIA) and a large panel of monoclonal antibodies (Mabs) derived for selected species of Leishmania, the isolates were characterized as members of the L. mexicana complex. These strains were different from those species infective to humans, in that they reacted with either the L.(L.) amazonensis or L. (L.) mexicana specific MAbs. Furthermore, this was also the case with cloned organisms from two stocks showing that they were not mixed populations of Leishmania (G Grimaldi et al. 1987 Am J Trop Med Hyg 36: 270-287). This group of distinct parasites presented, however, similar serodeme patterns in relation to the new recently described species of the Leishmania subgenus, L. (L.) forattinii (E Yoshida et al. 1993 Mem Inst Oswaldo Cruz 88: 397-406). The latter parasite was isolated from sylvatic reservoir hosts (Didelphis marsupialis aurita and P. iheringi denigratus) captured on regions endemic for American cutaneous leishmaniasis in Brazil, respectively in the municipalities of Conchas, São Paulo (E Yoshida et al. 1979 Rev Inst Med Trop S Paulo 21: 110-113) and Três Braços, Bahia (A Barreto et al. 1985 Rev Soc Bras Med Trop 18: 243-246). The general morphology and the growth characteristics in vitro of the new species were similar to those of other L. mexicana complex parasites. Here we have further studied the Espírito Santo isolates using additional techniques for the characterization of Leishmania. The molecular procedures used for typing the strains (isoenzyme electrophoresis, blot enzyme binding assay using MAbs, restriction endonuclease fragment patterns of k-DNA, and molecular karyotype analysis) have been described in detail in previous publications (Yoshida et al. 1979, 1983 loc. cit., Cupolillo et al. 1994 loc. cit., A Franco et al. 1997 Mem Inst Oswaldo Cruz 92: 63-68). Results of electrophoretic analysis were based on the following enzymatic loci, namely: glucose-6-phosphate dehydrogenase (G6PDH), malate dehydrogenase (MDH), isocitrate dehydrogenase (IDHNADP), 6-phosphogluconate dehydrogenase (6PGDH), nucleotidase (NH1 and NH2), glucose phosphate isomerase (GPI), phosphoglucomutase (PGM), proline dipeptidase (PEPD), leucine peptidase (PEP2), malic enzyme (ME), mannose phosphate isomerase (MPI), and aconitate hydratase (ACON). The enzyme profiles of the selected isolated signature MPRO/BR/82/RV228, which represents the poorly defined group of parasites from Espírito Santo, shared more electromorphs (bands of enzyme activity as revealed by electrophoresis) with L. (L.) forattinii and fewer with other members of the L. mexicana group (data not shown). The enzymes useful in distinguishing the new strain variant of L. (L.) forattinii are G6PDH, MDH, IDHNADP, PEPD, PEP2, and MPI. In addition, for most of the 13 enzyme loci examined, the profiles of this zymodeme (IOC-68) were different from those of all other species complexes of Leishmania. The taxonomical position of strain MPRO/BR/82/RV228 in relation to other selected species complexes of Leishmania ( Table) was defined by phenetic numerical analysis of the enzyme data (Cupolillo et al. 1994 loc. cit.). Affinities between zymodemes were calculated using the Jaccard´s similarity coefficient and were transformed into a phenogram, by the unweighted paired-group method using arithmetical avarage (UPGMA). As shown (Fig. 1), the two zymodemes classified as L. (L.) forattinii (IOC-11) and enzymatic variant (IOC-68) form distinct phenetic groups, clustered within the L. mexicana species complex. Fig. 1: phenogram showing the Jaccard´s coefficient of similarities between groups and/or species of Leishmania and the two strains (L67 and L182) of L. (L.) forattinii from Brazil. Western blot studies were performed using the L. mexicana complex specific Mabs (D McMahon-Pratt et al. 1985 J Immunol 134: 1935-1940) to further group/differentiate among these parasites. Antigens of each of the strains tested were prepared as promastigote homogenates containing protease inhibitors (Franco et al. loc. cit.). As shown (Fig. 2A), Mabs M9 and M11 recognized multiple molecular components with apparent relative mobility (Mr) values ranging from less than 14 to 200-kDa. Differences occurred between Leishmania species in either the Mr, intensity or number of components recognized by these Mabs. L. (L.) amazonensis fell into one subgroup (G1a). L. (L.) aristidesi showed as a distinct parasite (G1b), with an intermediary pattern between G1a and another subgroup (G1c), representing L. forattinii strains (which produced a similar profile, regardless of geographic distribution). In contrast, these antigenic determinant were not recognized in the remainder L. mexicana complex species analyzed. Although the banding patterns of these subgroups showed some similarities when tested with M4 and M6 (Fig. 2B), they were distinct from the other main group representing L. (L.) mexicana (G2). The main facts to emerge from this study are: (a) the presence of two major antigenically distinct groups in the L. mexicana complex; and (b) the apparent antigenic similarity within the group of the species (L.) amazonensis/L. (L.) garnhami, as well as the species L. (L.) mexicana/L. (L.) pifanoi. Fig. 2: western blot analyses of promastigotes homogenates of members of the Leishmania mexicana complex using monoclonal antibodies (A) M9 (XLV-2B5-H7) and M11 (XLV-1D11-E11), and (B) M4 (IX-1F9-D8) and M6 (LXVII-4C7-B8). The stocks codes of the strains analyzed are indicated below the lanes, and their origins are shown in the Table. Molecular weights are indicated in kDa besides the figure. Comparisons of kDNA restriction enzyme fragments profiles from stocks representing species complexes of New World Leishmania and the selected strains of L. (L.) forattinii from São Paulo (stock L67) and Espírito Santo (stock L182) were carried out by polyacrylamide gradient gel electrophoresis. Variation between the two strains of L. forattinii was demonstrated by their different schizodeme profiles with the enzymes Hinf I (Fig. 3A), Msp I (Fig. 3B), Hae III and Mbo I (data not shown). In addition, these profiles were distinct from those of all strains examined (Fig. 3). Fig. 3: acrylamide gradient (3.5-12%) gel electrophoresis comparison of kDNA fragment patterns, generated with the restriction enzyme Hinf I (A) and Msp I (B), among selected species complexes of Leishmania and the two strains (L67 and L182) of L. (L.) forattinii from Brazil. The identification of the Leishmania strains analyzed is given in the Table. Molecular weights of k-DNA fragments are indicated in basepairs (bp) besides the gel. In previous molecular karyotype analysis (Yoshida et al. 1983 loc. cit.), although L. forattinii and other species of the L. mexicana complex showed a specific number of size-concordant DNA molecules, clear karyotypic differences exist among these parasites. Moreover, chromosomes were polymorphic in both the number and size of bands between the two strains (L67 and L182) of L. forattinii analyzed in this study (data not shown). To date, no human infection with L. (L.) forattinii has been identified in the study areas. The species definitely associated with human and canine infections in Espírito Santo are (a) L. (Viannia) braziliensis, causing cutaneous ou mucosal leishmaniasis; and (b) L. (L.) chagasi, responsible for cases of visceral leishmaniasis (Grimaldi et al. 1989 loc. cit., A Falqueto et al. 1991 Mem Inst Oswaldo Cruz 86: 499-500). Here we have demonstrated that other leishmanial strains with similar characteristics to L. (L.) forattinii were recovered from Proechimys captured in the same region. Moreover, we have been investigating on the feeding habits of the phlebotomine sandflies in relation to human and Proechimys, in the forest area where the infected rodents were caught. Of 320 sandflies, 314 (98.1%) were identified as Lu. gasparviannai Martins, Godoy & Silva, 1962, indicating this species as the probable vector of the parasite among the rodents. No specimens of Lu. gasparviannai were found, however, among 355 phlebotomines caught feeding on humans (Falqueto et al. 1991 loc. cit.). These data suggest that the parasite here identified as a new enzymic variant of L. (L.) forattinii is not usually transmitted to humans. However, the pathogenicity of the new parasite was indistinguishable from other L. mexicana complex species, based on its virulence and development in laboratory animals (Yoshida et al. 1979, 1983 loc. cit.). Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98212a.jpg] [oc98212c.jpg] [oc98212d.jpg] [oc98212b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}