 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 93(6): 815-818 Prostaglandin A1 Inhibits Replication of Classical Swine Fever Virus Tânia Rosária Pereira Freitas, Lucio Ayres Caldas, Moacyr Alcoforado Rebello+ Instituto de Biofísica Carlos Chagas Filho and Departamento de Virologia,
Instituto de Microbiologia Received 27 October 1997; Accepted 13 July 1998
Code Number:OC98216
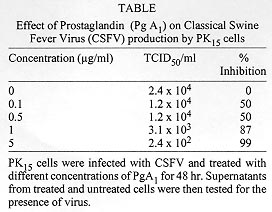
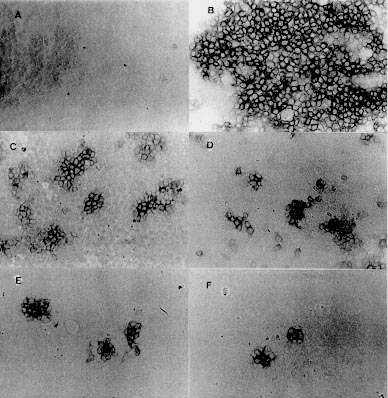
Prostaglandins (Pgs) have been shown to inhibit the replication of several DNA and RNA viruses. Here we report the effect of prostaglandin (PgA1) on the multiplication of a positive strand RNA virus, Classical Swine Fever Virus (CSFV) in PK15 cells. PgA1 was found to inhibit the multiplication of CSFV. At a concentration of 5 µg/ml, which was nontoxic to the cells, PgA1 inhibitis virus production in 99%. In PgA1 treated cells the size and number of characteristic Classical Swine Fever focus decreased in amount. Key words: Classical Swine Fever Virus - Prostaglandin - PK15 cell - virus replication Prostaglandins (Pgs) are a class of naturally occurring cyclic 20 carbon fatty acids, synthesized by most types of eukaryotic cells from polyunsaturated fatty acid precursors (Samuelson 1982). These compounds have been shown to function as microenvironmental hormones and intracellular signal mediators and to participate in the regulation of a large variety of physiological and pathological processes, including the immuno function (Goodwin & Webb 1980), interferon action (Pottathil et al. 1980), inflammation (Vane 1987) and virus replication (Santoro 1987). Pgs types A and J are characterized by the presence of an a, b unsaturated carbonyl group in the cyclopentane ring, and have been shown to inhibit the replication of several DNA and RNA viruses. However, the antiviral mechanism is not well understood and contrasting results have been reported for different virus-cell systems (Santoro et al. 1990). Classical Swine Fever (CSF) or Hog Cholera is a highly contagious viral disease of swines and can cause large economic losses. The infection is characterized by fever and hemorrahages and can lead to an acute, subacute, chronic, atypical, or inapparent manifestation. Acute CSF generally results in high morbidity and mortality, whereas infections with low-virulence virus can go unnoticed (Van Oirschot 1989). The Classical Swine Fever Virus (CSFV) is a member of Pestivirus genus, from Flaviviridae family (Horzineck et al. 1991). The Pestiviruses are enveloped, single-stranded positive polarity RNA with about 12,5 kb, and comprises a single large open reading frame. The messenger RNA is translated into a single polyprotein of about 3,898 amino acids which gives rise to mature proteins by proteolysis (Meyers et al. 1989). The structural proteins are located within the N-terminal one-third of the polyprotein (Stark et al. 1990). CSFV probably posseses four structural protein, three of which are glycosylated (Gp 44/48, Gp 33, and Gp 55). CSFV Gp 55 appears to be of major importance for neutralization of Pestivirus, since neutralizing antibodies are directed against this glycoprotein (Weiland et al. 1990). MATERIALS AND METHODS Cell culture and virus - PK15 cells were grown in Dulbecco, s Modified Eagle Medium (DMEM), supplemented with 10% tryptose phosphate broth and fetal bovine serum (Bovine Viral Diarrhea Virus - free), penicillin (500 U/ml), streptomycin (100 µg/ml) and amphotericin B (fungizone 2.5 µg/ml). CSFV Alfort-19 (A-19) strain, was provided by Laboratório de Referência Animal Pedro Leopoldo, MG, Brazil, and propagated in PK15 cells. PgA1 (Sigma Chemical Co.) was stored as a 100% ethanolic solution (1 mg/ml) and was diluted to the indicated concentrations in growth medium. Virus infection - A suspension of freshly trypsinised PK15 cells was dispensed in scintillation vials with growth medium and incubated in a humidified atmosphere of 5% CO2 and 95% air 37oC for 24 hr. The vials were voided of their medium and the monolayers were inoculated with approximately 100 TCID50/ml of a stock CSFV. After adsorption period of 1.5 hr at the same incubation conditions, the inoculum was eliminated and the infected cell culture were treated with different concentrations of PgA1. Virus assay - For the detection of infective VPSC a peroxidase linked assay (PLA) was used. The PLA was performed based in Afshar et al. (1989) and Office International des Epizooties - OIE Manual of Standards (1992) in microtitre plates and was standardized as described. 50 µl of double serial dilutions of virus samples were diluted in growth medium and distributed on the microtitre plate. Then, 150 µl of PK15 cells suspended in growth medium at a concentration of approximately 2 x 105 cells/ml were added to each well. After 48 hr, the monolayers were rinsed briefly with PBS, drained and fixed with 20% acetone in PBS. The monolayers were dried under a bench lamp during 3 hr and stored at -20oC. Prior to use, the monolayers were then rinsed with PBS, and overlaid with 50 µl of CSF-hyperimmune serum diluted 1:80 and incubated for 15 min at 37oC. The cells were rinsed three times with washing solution (8.85 g NaCl; 5 ml Tween 80 in 1L H2O destilated pH 7.2) and overlaid with 50 µl of a 1:2500 dilution of horseradish peroxidase, labeled rabbit anti-swine antibodies (Sigma, Chemical Co.). After an incubation of 10 min at 37oC, the wells were washed three times and filled with 50 µl of carbazol substrate solution (Sigma, Chemical Co.) prepared according to manufecter's recommendations. The plates were incubated for 30 min at 37oC in the dark and the cells were examined with an inverted light microscope. The VPSC foci was observed by the presence of reddish brown stained infected cells. RESULTS AND DISCUSSION Since antiviral effects of Pgs on CSFV have not been described, we first established in this paper the parameters involved in the protection of PK15 cells from this virus. Confluent monolayers of PK15 cells were infected with CSFV and treated with different concentrations of PgA1. Supernatants were collected 48 hr post-infection and virus titres were determined as described in Materials and Methods. Table shows that PgA1 inhibits virus yield with concentrations of 0.1 µg/ml being able to reduce virus yield by 50%. The infectious virus was progressively reduced resulting in 99% inhibition at 5 µg/ml PgA1. The antiviral effect clearly is not the result of PgA1, cytotoxicity as determined by either microscopic examination or by vital dye exclusion (data not shown). CSFV is mostly frequently isolated and growth in porcine kidney cell cultures. The virus replication is restricted to the cell cytoplasm and does not result in cytophathic effect (Laude 1987, Van Oirschot & Terpstra 1989). Porcine kidney cells such as PK15 cell line remain the most suitable system for its propagation. Due to the lack of cytophatic effect, accurate titration of the virus infectivity was based on the identification of microscopic fluorescent foci or by PLA (Jensen 1981, Edwards et al. 1991, Dahle et al. 1993, Freitas 1993). As we can see in Fig. 1B, in infected cells, there are stained spots which are the result of the peroxidase reaction, indicating the presence of virus, In PgA1 treated cells, the virus replication is impaired resulting in the appearance of small focus (Fig. 1C). In increased concentration of PgA1 (Figs 1D, E, F) the monolayers show a gradative decrease in the number and the size of the foci. Figure: Effect of Prostaglandin A1 on infectivity of Classical Swine Fever Virus (CSFV). PK15 cells were mock infected (A) or infected with CSFV (B, C, D, E, F) and maintained in growth medium (A, B) in the presence of PgA1 (C, D, E, F) at the concentrations of 0.1, 0.5, 1 and 5 µg/ml. After 48 hr the virus was detected in the monolayers as described in Materials and Methods. Photography was performed with an optical microscope. Magnification: X 200. Control cells (A), infected cells (B), infected cells treated with 0.1 µg/ml PgA1 (C) infected cells treated with 0.5 µg/ml PgA1 (D), infected cells treated with 1µg/ml PgA1 (E), infected cells treated with 5 µg/ml PgA1 (F). In the present study we have demonstrated that PgA1 inhibits the replication of CSFV in PK15 cells. To our knowledge this is first report concerning the effect of prostaglandins on the replication of virus from the Flaviviridae family. The antiviral activity of Pgs of the types A and J (Santoro 1987) has been described for several RNA and DNA viruses including orthomyxovi-ruses, picornaviruses, togaviruses, poxvirus, herpesviruses and retroviruses growing in different types of cells. Recently, we found that in PgA1-treated Aedes albopictus cells, the replication of Mayaro virus (Alphavirus genus, Togaviridae family) was also impaired (Barbosa & Rebello 1995). Even though an increasing amount of literature has now described the antiviral activity of Pgs, the mechanism by which these compounds can control virus replication is still mainly unknown. However, a relationship between heat-shock protein (HSP) synthesis (stress proteins) and virus replications has been described by several authors. The possibility that HSP could be part of an intracellular defense strategy against viruses is fascinating. Heat shock protein synthesis by Pgs could also be linked to alterations in the glycosylation process and in the control of cell proliferation (Santoro et al. 1990). Due to the many cellular functions of HSP and to the many levels of interactions described above, the relationship between viruses and heat schock proteins appears to be a complex phenomenon. Data in the literature are insuficient to suggest a mechanism for the effect of PgA1. Holbrook et al. (1992), studying the effect of PgA2 on Hela cells demonstrated that PgA2 induces high levels of HSP70 mRNA, which results from an increase in the rate of HSP70 genes transcription. Pica et al (1993) studying the action of PgJ2 in the replication of vesicular stomatitis virus found that the induction of HSP synthesis is accompanied by a decrease in [3H] glucosamine incorporation into the virus glycoprotein G, at concentrations which do not alter glucosamine uptake by the cells, suggesting that a impairment in virus protein glycosylation could be responsible for the antiviral activity. This work was supported by a research grant from CNPq, FINEP and FUJB. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98216a.jpg] [oc98216b.jpg] |
| |||||||||

{kind=link}
{kind=link}