 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
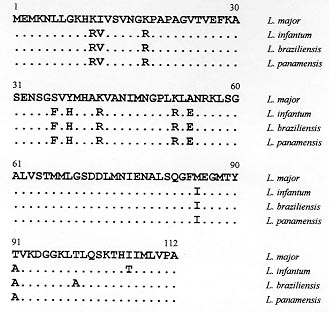
Vol. 93(6): 819-821 RESEARCH NOTE The Metacyclic Stage-expressed Meta-1 Gene is Conserved between Old and New World Leishmania Species Christof Berberich+, Marcel Marín++, José R Ramírez, Carlos Muskus+++, Ivan D Vélez Programa de Estudio y Control de Enfermedades Tropicales, Sección de
Inmunología y Biología Molecular, Facultad de Medicina, Universidad de
Antioquia, A.A. 1226, Medellín, Colombia Received 13 April 1998; Accepted 18 June 1998
Code Number:OC98217
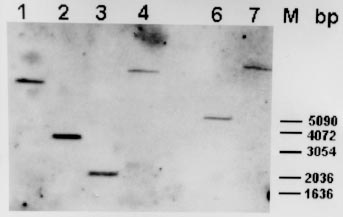
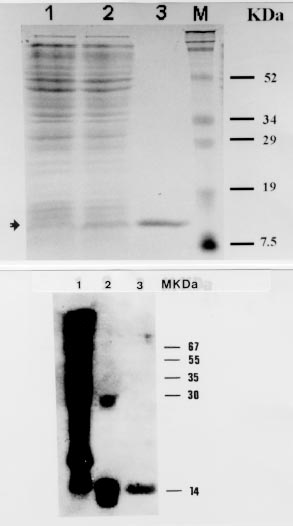
Key words: leishmaniasis - Leishmania infantum - Leishmania (Viannia) braziliensis -Leíshmania (Viannia) panamensis - meta-1 gene -metacyclogenesis - antigenicity RESEARCH NOTE Leishmaniasis is an endemic disease in large parts of the world including Central and South America. The most prevalent species in Colombia belong to the complex Leishmania (Viannia) braziliensis and are responsible for the majority of cases of cutaneous, mucocutaneous and dissiminated cutaneous leishmaniasis (A Corredor 1990 Am J Trop Med Hyg 42: 206-214). Cases of visceral leishmaniasis in Colombia, in contrast, are always caused by L. (Leishmania) infantum/chagasi (A Corredor 1989 Am J Trop Med Hyg 5: 480-485). Within the promastigote life-cycle stage in the insect alimentary tract, Leishmania sequentially develops from proliferating, non-infective (procyclic) to non-dividing, infective (metacyclic) forms. Despite of substantial progress in the understanding of some biochemical aspects of this differentiation process, the molecular mechanisms underlying metacyclogenesis remain to be elucidated. In an attempt to dissect the signals inducing the metacyclic differentiation step, genes were recently identified to be predominantly or exclusively expressed in metacyclic promastigotes compared with proliferating forms, including the ORFs mat-l (T Brodin 1992 Mol Biochem Parasitol 52: 241-250), gene B (H Flinn 1994 Mol Biochem Parasitol 65: 259-270) and meta-l (F Nourbakhsh 1996 Mol Biochem Parasitol 76: 201-213). Given the unique expression pattern of these genes during metacyclogenesis it can also be hypothezised that these proteins are associated with Leishmania infectivity, thus rendering them interesting molecules for detailed studies of their gene functions. To date, only the L. major meta-1 gene has been partially characterized, and it has been shown that it codes for a putative protein of 112 amino acids, at least 20-fold upregulated during metacyclogen-esis (Nourbakhsh loc. cit.). The present work was undertaken in order to isolate the respective meta-1 ORFs in L. infantum and species of the subgenus Viannia, including L. braziliensis and L. panamensis, in order to determine the genomic organization of the meta-1 locus and in order to express meta-1 as recombinant protein in Escherichia coli for further antigenicity analysis using sera from leishmaniasis patients. For that purpose two 30-bp oligonucleotide primers (sense- 5' CGCGGATCCATGGAGATGAAAAA CTTGCTT 3', including the restriction site Bam HI; antisense: 5' CCCAAGCTTCGCAGGAAC AAGCATGATGAT 3', including the restriction site Hind III) which sequences were based on the recently published meta-1 gene from L. major (Nourbakhsh loc. cit.) were synthesized to PCR-amplify the meta-l coding region using the genomic DNA of L. infantum (MHOM/FR/78/LEM75/MON1), L. braziliensis (MHOM/BR/75/M2903) and L. panamensis (MHOM/PA/71/LS94) as template. Under stringent annealing conditions for the PCR-reactions (annealing temperature 58oC) only one DNA amplification product corresponding to a 336-bp fragment was obtained. The PCR-fragment was subsequently cloned in the polylinker of the pBluescript vector for sequencing of both DNA strands. Fig. 1 shows the deduced amino acid sequence of the meta-1 coding region of L. infantum, L. braziliensis and L. panamensis, marking the differences to the recently published L. major sequence (Nourbakhsh loc. cit.). The meta-l open reading frames are quite conserved in all species (about 90% with respect to L. major). As expected, only one amino acid substitution within the subgenus Viannia was observed. Remarkably, most of the amino acid modifications with respect to L. major are found in L. infantum as well as in the Viannia species (only two modifications between L. infantum and L. braziliensis and one substitution between L. infantum and L. panamensis). Thus, the meta-1 sequences are more conserved between these three species than when compared to the L. major sequence. The meta-1 protein of L. infantum as deduced from the respective amino acid sequence has a molecular mass of 12 kDa and an isoelectric point of 9.54. The molecular analysis of the putative secondary structure using the Garnier-Robsen approximation (J Garnier 1978 J Mol Biol 120: 97-120) shows a relatively high percentage of alpha-helix and beta-sheet regions with only two short stretches of coil-structures at the N-terminus and in the central protein segment (data not shown). Fig. l: complete amino acid sequence of the meta-1 ORF in Leishmania infantum, L. braziliensis and L. panamensis. Dots indicate identical amino acids, and differences with respect to the L. major protein are given by the corresponding letters. In order to determine the genomic arrangement of the meta-1 gene locus in L. infantum, we performed a southern blot analysis of restriction-digested DNA isolated from L. infantum pro-mastigotes. As shown in Fig. 2, only one hybridizing DNA-fragment can be observed, independently of the type of restriction enzyme used, including Pst 1 that cuts within the meta-1 coding sequence. Therefore, our results point to the existence of a single gene copy of meta-1 in L. infantum and confirm similar observations in the L. major gene locus (Nourbakhsh loc.cit.). Fig. 2: southern blot analysis of the genomic organization of the meta-1 locus in Leishmania infantum 2 mg of genomic DNA were digested with various restriction enzymes (1: Bam HI, 2: Nco 1, 3: Pst 1, 4: Xho 1, 6: Sal 1, 7: Bcl l), blotted to a nylon membrane and hybridized to the 336 bp PCR-amplified meta-l DNA-fragment used as probe. The length of the DNA-fragments produced by the molecular weight marker are indicated. Since the meta-l gene appears highly up-regulated in infective promastigotes, we were interested in determining whether this protein may act as an antigenic component during natural course of leishmaniasis in infected humans. The entire coding regions of the L. infantum and L. panamensis meta-l were therefore introduced in frame into the bacterial expression vector pQE 30. Transformation of E. coli with these constructs resulted in high level-expression of the recombinant, His-tagged meta-l proteins and allowed the purification of the proteins by passage of the whole bacterial extract through Ni-NTA affinity columns. The purity and yield of the recombinant proteins was analyzed in SDS-PAGE gels (Fig. 3a). The L. infantum protein was subsequently transferred to nitrocellulose filters and incubated with a pool of two sera obtained from human visceral leishmaniasis patients living in an endemic area for the visceral form of the disease. The western blot shown in Fig. 3b demonstrates that the recombinant protein was efficiently recognized by antibodies present in the sera. As positive control we used the recombinant KMP-11 protein, recently described as a strong antigenic component of the Leishmania parasite (C Berberich 1997 Exp Parasitol 85: 105-108). When incubating the recombinant L. panamensis meta-1 counterpart with sera from Colombian patients suffering from cutaneous and mucocutaneous leishmaniasis we could also observe in ELISA studies the presence of anti-meta-l antibodies, although to a lesser extent than in visceral leishmaniasis sera (percentage of positive sera, approx. 30%; data not shown). Sera from healthy, non-Leishmania infected individuals did not show any reactivity against the recombinant meta- l. Fig. 3: recombinant meta-1 expression in Escherichia coli and western blot analysis. A: SDS/PAGE-analysis of the bacteria expression and purification of the Leishmania infantum meta-1 protein. l: uninduced bacteria, 2: induced bacteria (2 mM IPTG), 3: recombinant His-tagged meta-1 after affinity purification on Ni-NTA. The bands produced by the protein molecular weight marker are indicated. B: western blot analysis of the recombinant L. infantum meta-l. 5 mg of total protein lysate of L. infantum promastigotes (1), 500 ng of the recombinant KMP-11 (2) and 500 ng of the recombinant meta-1 (3) were run on a 12% SDS-PAGE, blotted to a nitrocellulose membrane and subsequently incubated with a pool of two human visceral leishmaniasis sera (dilution l: 100). In conclusion, we show evidence that the meta-1 gene is conserved in the genus Leishmania, independently of the geographic distribution of individual species and clinical symptoms they cause. The protein is an antigenic component of the parasite, although the prevalence of antibodies, in particular in cutaneous and mucocutaneous leishmaniasis sera, may be in some cases too low in order to consider meta-1 as a highly sensitive marker for diagnosis of infection. Given the unique expression pattern of meta-1 in infective metacyclic promastigote forms and the single-copy gene structure of the genomic locus, we believe that this protein may be an ideal candidate for studies on the gene-function relationship using the gene-knock out approach. We are also currently focusing on the hypothesis that this metacyclic protein may be involved in evasion mechanisms of the incoming parasite from macrophage-mediated killing mechanisms. Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98217c.jpg] [oc98217b.jpg] [oc98217a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}