 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 93(6): 861-864 The Effects of a DNA Virus Infection on the Reproductive Potential of Female Tsetse Flies, Glossina morsitans centralis and Glossina morsitans morsitans (Diptera: Glossinidae) Rosemary C Sang/+, Walter GZO Jura*, Leonard H Otieno*,Richard W Mwangi** Virus Research Centre, Kenya Medical Research Institute, P.O. Box 54628, Nairobi,
Kenya Received 5 May 1998; Accepted 22 July 1998
Code Number:OC98225
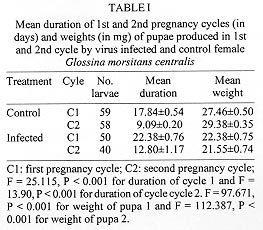
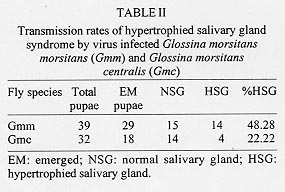
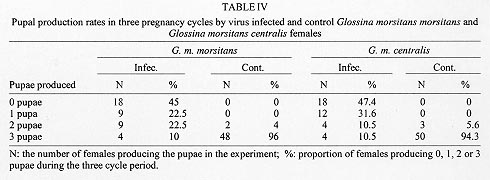
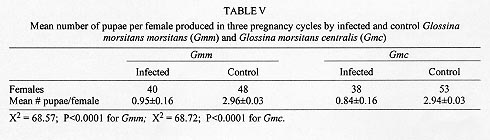
Reproductive anomalies associated with the tsetse DNA virus infection in the female tsetse hosts, Glossina morsitans centralis Machado and Glossina morsitans morsitans Westwood, inoculated with the virus during the 3rd instar larval stage were studied and the data compared to those obtained from the control females injected with sterile physiological saline. Virus infected flies had significantly longer first and second pregnancy cycles (P<0.0001) and produced pupae that were of significantly less weight in milligrams (P<0.0001) compared to controls. Transmission of the virus to progeny was not absolute and only 21% of G. m. centralis and 48% of G. m. morsitans first progeny flies from infected females developed salivary gland hypertrophy as a result of transmission from mother to progeny. The virus infected females produced significantly fewere pupae compared to the controls during the experimental period (P<0.00001). Key words: DNA virus - tsetse fly - Glossina morsitans centralis - Glossina morsitans morsitans - reproductive potential - pregnancy cycle - pupal weights - transovarial transmission The tsetse virus causing salivary gland hypertrophy and male sterility has been shown to infect some tsetse fly species naturally in the wild including Glossina pallidipes and Glossina morsitans morsitans (Whitnal 1934, Jaenson 1978, Otieno et al. 1980, Ellis & Maudlin 1987). Subsequently it has been partially characterized as a double stranded DNA virus (Odindo et al. 1986). A method of infecting tsetse flies in the laboratory was developed by Jura et al. (1993), thus facilitating laboratory experimentation on this virus to determine how it affects its hosts. Tsetse flies are reproductively peculiar because the females give birth to larvae, nourished within the mother by a secretion from the highly modified uterine milk gland (Tobe & Langley 1978). The first oocyte is ovulated about nine days after emergence of the female (Saunders 1970) and is fertilised as it enters the uterus. Embryogenesis takes place for three days after which the larva hatches and goes through three larval instars in the uterus. It is born as a full term 3rd instar larva at the end of a first pregnancy cycle which takes 18 days in the laboratory conditions while subsequent cycles take 9 to 10 days (Langley 1977). The larva pupates in 15 to 30 min and the adult fly emerges from the pupa 30 days later. Tsetse thus have a low reproductive potential in comparison to other insects as only a small number of developed larvae can be produced in the life of a female. The fecundity of tsetse is thus highly dependent on the survival rate and the duration of interlarval period. Jura et al. (1988) examined the ovarioles of virus infected G. pallidipes and observed that the majority of the germaria were affected by degeneration and severe necrosis and virus particles were seen within the germarial cells suggesting that this virus may be transmitted transovarially to the progeny by an infected female. The degenerative changes may affect the development of the larva resulting from such an egg. Sang et al. (1997) demonstrated that flies infected by this virus had difficulty in feeding and this affected their survival significantly. These factors would probably influence the reproductive performance of the infected female fly. Evaluation of the potential of this virus as a biocontrol agent in the control of tsetse flies includes obtaining information on the effects of the virus on the female reproductive potential as well. The specific objectives of this study were to investigate the effects of this virus on the duration of the pregnancy cycles, pupal productivity and their weights. MATERIALS AND METHODS Virus material - G. pallidipes were trapped at the Ruma National Park in the Lambwe Valley Game Reserve in Western Kenya. Flies were dissected in the laboratory and any hypertrophied salivary glands retrieved and frozen in sterile physiological saline. Fifteen pairs of these were later homogenised in 0.5 ml of sterile saline centrifuged at 3,000 rpm for 15 min at 4oC. The supernatant was used to inoculate experimental flies. Production of virus infected and control flies - Freshly larviposited larvae of G. m. centralis or G.m. morsitans were obtained from the insectary where tsetse are reared at 25oC and 75-80% relative humidity. The larvae were inoculated with 1 ml of virus suspension prepared as above using the method developed by Jura et al. (1993). This technique confers 100% infection in treated flies. Control flies were inoculated with 1 ml of sterile saline. After the larvae pupated they were incubated under the insectary conditions for approximately 30 days to emerge. At emergence the sexes were caged separately in PVC fly holding cages. Flies were always fed on a live rabbit. Effects of virus infection on pregnancy cycle, pupal production - Virus-infected 2-day old G.m. centralis females were fed on rabbit blood and after few hours mated to 8 day-old males obtained from the insectary, where they are fed every other day also on rabbit blood. The males were removed after 24 hr of mating and destroyed. The females were tranferred to single fly holding plastic vials measuring 5 x 2.5 cm with a nylon netting on one end to allow feeding, kept under insectary conditions and fed every other day. A group of control flies were treated similarly. Flies were observed for two pregnancy cycles and the duration of the cycles, pupal production, weights of pupae and their emergence were recorded. Transmission rate of the virus from the mother to progeny - Both G. m. morsitans and G. m. centralis were used in these experiments. Females of these species emerging from both control and virus infected pupae were fed on rabbit blood on day 2 and then mated to 8 day-old males as in the previous experiment. The flies were caged singly in single-fly holding plastic vials, fed regularly and checked daily for pupal production for three pregnancy cycles. The pupae from each fly were caged separately in single fly holding tubes and upon emergence they were dissected and examined for salivary gland hypertrophy. At the end of the three pregnancy cycles, the experimental females were dissected to check for insemination and to confirm salivary gland hypertrophy in the virus infected flies. RESULTS Effects of virus infection on pregnancy cycle and pupal weights - It was observed that the durations of the first and the second pregnancy cycles in virus infected flies were significantly longer than in the controls and the pupae produced by virus infected females were of significantly lower weights compared to those produced by control females (P<0.0001, Table I). Virus transmission rates to progeny flies by virus infected females - It was observed that an infected female did not always transmit the virus to all her progeny. Only 21% of the progeny from G. m. centralis and 48% of the progeny from G. m. morsitans developed the syndrome (Table II). Where an infected fly was able to produce more than one puparium, in most cases it either produced all infected progeny or all normal progeny except two flies (nos. 14 and 18, see Table III) which produced mixed progeny (one virus infected and one uninfected). The rest of the flies in the experimental group produced only one or no larvae at all and in few cases the larvae produced failed to pupate. Pupal productivity - Table IV demonstrates that 45% of the female G. m. morsitans and 47% of G. m. centralis did not produce any larvae at all through the experimental period although they were inseminated. Some of the females either died before they larviposited or failed to larviposit over the experimental period. Only 10% the virus infected G. m. morsitans and G. m. centralis females produced three larvae during the experiment. Among the controls of both species, over 90% of the females produced three larvae during the experimental period and as demonstrated in Table V, infected females produced significantly more pupae per female compared to the controls. DISCUSSION These results reveal that the tsetse virus infection has detrimental effects on the female reproductive performance, lengthening the interlarval periods and reducing the pupal production and their weights. The reduction in the weights of pupae of infected flies can be attributed partly to the presence of necrotic lesions in the germaria and viral particles detected in the germarial cell nuclei (Jura et al. 1988). The presence of virogenic stromata in these cells is an indication of replicating sites of the virus. Replication and assembly of virions in cells provides a stress on the affected organ as the cell macromolecular synthetic machinery is switched over to the synthesis of viral protein. The replication of the virus in the ovarioles could have an impact on the nutritive requirements and other developmental factors which could lead to delays in larval development or lack of it. This could also partly explain the increased interlarval periods. The replication of the virus and the pathological changes observed in milk glands of virus infected females (Sang et al. 1996) may also lead to the poor nutritional provision to the larva in utero and could also explain the reduced pupal weights and lengthened interlarval periods. Other observations (Sang et al. 1997) have shown that infected flies experienced difficulty in feeding and in some cases the digestion was greatly impaired due to lesions occurring in sections of the midgut. This could also affect the development of the larvae in utero as the larva depends on the nutritive provision of the mother for its development. Lesions occurring in the midgut would result in impaired digestion and absorption and this would lead to undernourishment of the infected female and hence the larva in utero. It has been reported in some laboratory studies that the amount of blood embibed during interlarval period is a major factor determining the reproductive performance of tsetse (Langley & Stafford 1990, Gaston & Randolph 1993). Saunders (1970) showed that factors other than the temperature can alter the interlarval period and also suggested that the milk gland secretion is perhaps initiated by correctly timed blood meals and in the absence of such blood meals the secretion of the milk is delayed and the larva becomes undernourished with consequent retardation in its growth rate. Results of this study further demonstrate that although a female may show salivary gland hypertrophy, it may fail to transmit the infection to the progeny. Jaenson (1978, 1986) working on field-collected and laboratory reared G. pallidipes recorded that infected females, in general, produced infected progeny. However, Jura et al. (1988) demonstrated that only occasional virus particles were found within the ooplasm and the affected oocytes appeared intact without pathological changes. Sang et al. (1996) demonstrated the presence of the virus in milk gland cells with virogenic stroma in the cell nuclei and suggested that the virus could be transmitted through milk secretion to the larva in utero. This mode of transmission probably occurs when sufficient virus particles are ingested with the milk. These effects of the virus on the female reproductive potential partly explain the reported low incidence of the virus infection in nature which ranges between 0.5% to 5%, (Otieno et al. 1980, Ellis et al. 1987, Gouteaux 1987). It is possible that the establishment of this virus in a tsetse infested area could help to regulate the tsetse population by acting as a control agent. Extensive field studies are required to determine the role of the DNA virus in the regulation of tsetse populations in nature. ACKNOWLEDGEMENTS. To Dr Peter Tukei, Director of the Virus Research Centre, for granting permission for this work to be done and to James O Adino for technical assistance. This paper has been published with the approval of the Director of the Kenya Medical Research Institute. REFERENCES
Copyright 1998 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc98225c.jpg] [oc98225a.jpg] [oc98225d.jpg] [oc98225b.jpg] [oc98225e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}