 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
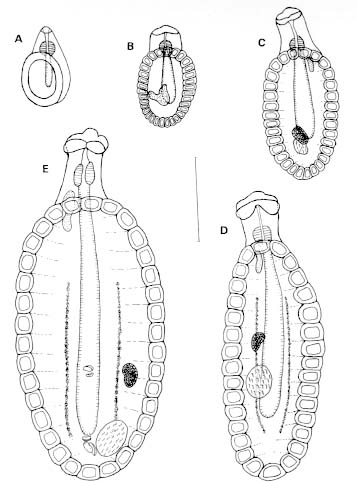
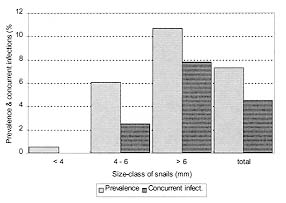
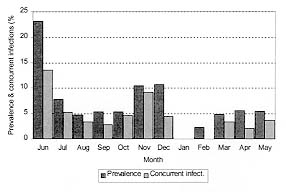
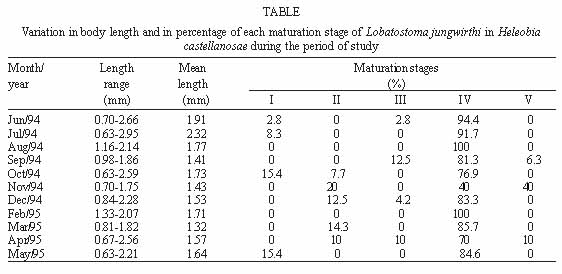
Vol. 94(1): 31-35 Some Aspects of the Development of Lobatostoma jungwirthi Kritscher, 1974 (Aspidogastrea) in Snails and Cichlid Fishes from Buenos Aires, Argentina María Inés Zylber+, Margarita Ostrowski de Núñez Laboratorio de Helmintología, Departamento de Ciencias Biológicas,
Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria,
Pabellón II, (1428) Buenos Aires, Argentina Received 20 May 1998; Accepted 22 October 1998
Code Number:OC99010
The molluscs Heleobia castellanosae and Ampullaria canaliculata and cichlid fishes were collected from an artificial pond at Tres de Febrero Park (Buenos Aires city), between June 1994 and May 1995. One hundred and eighty of 2,467 H. castellanosae examined were infected with pre-adults of Lobatostoma jungwirthi, 112 of them in concurrent infections with digeneans. L. jungwirthi was significantly more prevalent in larger snails, especially those infected with larval digeneans, but the prevalence of infection did not fluctuate significantly throughout the year. A. canaliculata and cichlids smaller than 10 cm were not infected, but larger Cichlasoma facetum harboured adults of L. jungwirthi in their intestines. Experimental infections of C. facetum and Gymnogeophagus meridionalis with pre-adult aspidogastreans obtained from H. castellanosae were successful. The development of L. jungwirthi in the snail host involves five arbitrary growth phases. Its life-cycle is heteroxenous, similar to that described for L. manteri, with H. castellanosae as the intermediate host and cichlid fishes as the definitive host in this pond. Key words: Aspidogastrea - Lobatostoma jungwirthi - Heleobia castellanosae - concurrent infections - prevalence of infection - seasonality - Buenos Aires - Argentina In one of the artificial ponds at Tres de Febrero Park (Buenos Aires city), Szidat (1971) found a species of the genus Lobatostoma Eckmann, 1932 (Aspidogastrea) parasitizing the gonad of the prosobranch snail Heleobia castellanosae (Gaillard, 1974) Davis, Mazurkiewicz and Mandracchia, 1982 and considered it identical to L. pacificum Manter, 1940. Since L. pacificum parasitizes marine fishes, its taxonomic status seems to be quite uncertain and needs to be reconsidered. In this work, the species of Lobatostoma from the same snail and pond as that found by Szidat is identified as L. jungwirthi Kritscher, 1974. L. jungwirthi was first described parasitizing the freshwater cichlid fish Geophagus brachyurus Cope from Rio do Sinos, Brazil. In 1984, Lunaschi redescribed this species in Cichlasoma facetum (Jenyns) from Buenos Aires Province. This is the only member in the genus which parasitizes freshwater fishes. Operculate snail species serve as intermediate hosts not only for digenetic but also for various aspidogastrean trematodes (Rohde 1972, Hendrix et al. 1985, Huehner 1987). Rohde and Sandland (1973) reported Lobatostoma manteri Rohde, 1973 to occur more frequently in a prosobranch snail naturally infected with digenean parthenitae. Huehner (1975, 1987) experimentally demonstrated that Aspidogaster conchicola von Baer could successfully establish itself and develop in the gastropod Elimia livescens with pre-existing digenetic infections, and that these aspidogastrean-digenean double infections could occur seasonally in nature. Prevalences of single and concurrent infections of L. jungwirthi in H. castellanosae from Buenos Aires city are reported herein. Since its discovery, the morphology of L. jungwirthi has been thoroughly studied, but not so its development and transmission. Aspidogastrean life-cycles have been worked out for Aspidogaster indicum Dayal (see Rai 1964), Multicotyle purvisi Dawes (see Rohde 1972) and A. conchicola (see Huehner & Etges 1972, 1977, Bakker & Davids 1973, Rohde 1994). Among the species of the genus Lobatostoma, only the life-cycle of the Australian L. manteri from the marine teleost Trachinotus blochi (Lacépède) is known (Rohde 1973, 1994). The present study analyzes L. jungwirthi's development in its molluscan host and elucidates its life-cycle. MATERIALS AND METHODS Monthly samples of H. castellanosae (Prosobranchia: Hydrobiidae) were collected using nets from a pond at Tres de Febrero Park between June 1994 and May 1995. A. canaliculata (Prosobranchia: Ampullariidae) and the cichlid fishes C. facetum and G. meridionalis were irregularly captured in the same pond. The snails were measured and dissected in the laboratory. Lobatostoma specimens were collected from the gonads of their molluscan hosts, washed in 0.3‰ NaCl saline solution, fixed with Raillet-Henry's fixative (8‰ NaCl saline solution, formalin, acetic acid), stained with alcoholic-hydrochloric carmine (Langeron 1949), cleared in creosote and mounted in Canada balsam. The following maturation stages were arbitrarily established for the analysis of the aspidogastreans in the snail host, according to their size and morphology. Class I: very small, up to 0.8 mm long; forebody (body region anterior to ventral sucker) short, without head lobes surrounding mouth; adhesive ventral sucker subcylindrical and not yet alveolated; sexual organs and vitelline glands inconspicuous (Fig. 1A). Class II: small, not exceeding 1 mm long; forebody short, still lacking head lobes; ventral sucker subcylindrical, with developing marginal alveoli; testis and ovary absent or starting to develop, vitelline follicles still inconspicuous (Fig. 1B). Class III: medium-sized, up to 1.25 mm long; forebody of medium size, with head lobes still absent or starting to develop; ventral sucker oval, with fully-formed marginal alveoli and developing median alveoli; longitudinal median ridge appears dividing median alveoli; sexual organs and vitelline follicles rudimentary (Fig. 1C). Class IV (pre-adults): large, (1-3) mm long; forebody large, with head lobes completely developed; ventral disc oval, with complete alveolation; genitalia and vitelline follicles fully developed (Fig. 1D). Class V (pre-adults): as class IV, but with presence of 1-8 eggs (Fig. 1E). Fig. 1: maturation stages of Lobatostoma jungwirthi: A: class I; B: class II; C: class III; D: class IV; E: class V. Bar = 500 µm. The fishes C. facetum and G. meridionalis, captured in a pond where H. castellanosae was absent, were experimentally infected several times with aspidogastrean pre-adults. They where manually fed with the dissected bodies of the parasitized snails. A chi-square test including Yate's correction with a two-tailed P value was carried out to analyze differences between prevalence of infection of L. jungwirthi in singly and concurrently infected H. castellanosae. Kruskal-Wallis non-parametric ANOVA tests were employed to evaluate the statistical significance of the associations between prevalence of infection, snail size and month, both for single and double infections. The level of significance was set at P<0.05 (Rohlf & Sokal 1981). Unless otherwise stated, all measurements are given in millimetres. We used "prevalence of infection" as defined by Bush et al. (1997). RESULTS The parasites obtained from H. castellanosae and from experimentally infected C. facetum were identified as L. jungwirthi Kritscher, 1974, as redescribed by Lunaschi (1984). L. jungwirthi can be distinguished by the absence of a tail overlapping the ventral disc posteriorly, the testis located in the median third of the body, a small, ovoid cirrus-pouch and the presence of 30-40 marginal alveoli on the ventral disc. Of 2,467 H. castellanosae examined, 180 (7.3%) had their gonads infected with one, rarely two or more, immature specimens of L. jungwirthi. Prevalence of infection varied significantly, depending on snail size: L. jungwirthi was more prevalent in larger snails (K-W=21.389, P<0.0001) (Fig. 2). Although it showed a peak in the month of June (23.1%) (Fig. 3), the prevalence did not display any significant changes throughout the year. Fig. 2: Lobatostoma jungwirthi and aspidogastrean-digenean concurrent infections in Heleobia castellanosae in different host size-classes: prevalence of infection. Fig. 3: Lobatostoma jungwirthi and aspidogastrean-digenean concurrent infections in Heleobia castellanosae: monthly prevalence of infection 1994-1995. L. jungwirthi exhibited significant variations in prevalence of infection between singly and concurrently infected snails: it co-occurred with digeneans to produce 112 (4.5%) double infections, while 68 snails lacked digenean larval stages (2.8%) (c2=12.519, P=0.0004). Aspidogastrean-digenean concurrent infections were significantly more common in larger snails (K-W=18.628, P<0.0001) (Fig. 2), but they did not vary significantly throughout the year (Fig. 3). Worm body lengths ranged from 0.63 to 2.95, with mean values ranging between 1.32 in March 1995 and 2.32 in July 1994 (Table). The smallest individuals (class I) were observed irregularly between May and October, while those belonging to class II appeared from October to April. Class III was observed in April, June, September and December, whereas the largest individuals (class IV) were always prevalent (Table). Only four specimens belonging to class V were found, one carrying a single egg (April), another with two eggs (September) and two with one and eight eggs within the uterus (November) (Table). Eighteen A. canaliculata plus 33 G. meri-dionalis and C. facetum (<10 cm long) captured in the pond were not infected with L. jungwirthi. Two C. facetum (11 and 12.5 cm long) harboured in their intestines ten and one specimens, respectively; the latter being mature and of great size (3 mm). One of seven experimentally infected G. meridionalis (4.5 cm long) carried six L. jungwirthi in the rectum, one of them showing incipient egg production 24 hr after the last infection. Six of eight experimentally infected C. facetum (8- 9.5 cm long) yielded 16 specimens of L. jungwirthi, three of them with eggs after 96 and 72 hr following the last infection. DISCUSSION At present, nine species are considered as valid members of the genus Lobatostoma: L. ringens (Linton, 1905); L. kemostoma (MacCallum & MacCallum, 1913); L. pacificum Manter, 1940; L. manteri Rohde, 1973; L. jungwirthi Kritscher, 1974; L. platense Mañé-Garzón & Holcman-Spector, 1976; L. hanumanthai Narashimhulu & Madhavi, 1980; L. anisotremum Oliva & Carbajal, 1984, and L. veranoi Oliva & Luque, 1989 as cited by Oliva and Luque (1989) and Rohde (1994). L. ringens, L. pacificum, L. jungwirthi, L. platense, L. anisotremum and L. veranoi are South American species. Only L. jungwirthi parasitizes freshwater, rather than marine fishes. Except for the Australian L. manteri, whose intermediate host is a mollusc (Rohde 1973), the life cycles of the rest of the species remain unknown. The positive correlation between larger sizes of H. castellanosae and a higher probability of trematode infection may simply result from the fact that older and larger snails have a greater exposure to the parasite's eggs because they eat and are more mobile than smaller individuals, as observed by Huehner (1987) in the case of the host of A. conchicola. The lack of seasonality in terms of prevalence may imply that cycles other than seasonal ones may be involved, since individuals belonging to all maturation stages, except class V, appear throughout the year. The level of aspidogastrean-digenean concurrent infection appears to follow the aspidogastrean prevalence pattern with regard to snail size and seasonality. Rohde and Sandland (1973) reported a significant tendency for L. manteri to occur more frequently in snails with digenean infections and suggested that a reduced resistance to a second infection may be responsible for this pattern. Huehner (1987) reported the same tendency for A. conchicola in the gastropod E. livescens and suggested that some digeneans may produce beneficial effects on concurrent aspidogastrean infections. These results are consistent with the present results for L. jungwirthi in snails with digenean infections. According to Huehner (1987), some findings demonstrate that digenean infections significantly increase the probability of aspidogastrean infection and vice versa. The mechanisms underlying this relationship, either physiological (reduced resistance) or behavioural (i.e., increased feeding or exposure to parasite eggs or miracidia), remain unknown. Rohde (1972, 1994) had emphasised the low degree of host specificity of aspidogastreans, both for the mollusc and the vertebrate hosts, with possible exceptions. For example, three snail species serve as hosts for L. manteri at the Great Barrier Reef, Australia. In the study area, L. jungwirthi was exclusively found in the mollusc H. castellanosae. No definite conclusion on host specificity in vertebrates can be formulated, since only a few specimens of all of the potential hosts from one area have been examined. Infection experiments are also inconclusive, because unnatural conditions may influence the results (abnormal diet, stress, etc.). According to Rohde (1973), experimental data should be used with caution when formulating statements on the host specificity of parasites. The life-cycle of L. jungwirthi is heteroxenous, very similar to that described by Rohde (1973, 1994) for L. manteri which, according to the author, is the simplest obligatory two-host life-cycle known for trematodes. In his study, Rohde showed, for the first time for the Aspidogastrea, that a vertebrate has become an obligatory host in which sexual maturity is reached: L. manteri is not capable of completing its cycle in snails. L. jungwirthi also needs a vertebrate host to complete its life-cycle, at least in the study area and for the examined hosts. REFERENCES Bakker KE, Davids C 1973. Notes on the life history of Aspidogaster conchicola Baer, 1826 (Trematoda; Aspidogastridae). J Helminthol 47: 269-276. Bush AO, Lafferty KD, Lotz JM, Shostak AW 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83: 575-583. Hendrix SS, Vidrine MF, Hartenstine RH 1985. A list of records of freshwater aspidogastrids (Trematoda) and their hosts in North America. Proc Helminthol Soc Wash 52: 289-296. Huehner MK 1975. Studies on the biology of Aspidogaster conchicola von Baer, 1827, Cotylaspis insignis Leidy, 1858 and Cotylogasteroides barrowi Huehner and Etges, 1972. Dissertation Abstracts International 36B: 1611-1612. Huehner MK 1987. Aspidogastrid and digenetic trematode single and double infections in the gastropod, Elimia livescens, from the upper Cuyahoga River. Proc Helminthol Soc Wash 54: 200-203. Huehner MK, Etges FJ 1972. Experimental transmission of Aspidogaster conchicola von Baer, 1827. J Parasitol 58: 109. Huehner MK, Etges FJ 1977. The life cycle and development of Aspidogaster conchicola in the snails, Viviparus malleatus and Goniobasis livescens. J Parasitol 63: 669-674. Kritscher E 1974. Lobatostoma jungwirthi nov. spec. (Aspidocotylea, Aspidogastridae) aus Geophagus brachyurus Cope, 1894 (Pisc., Cichlidae). Ann Naturhistor Mus Wien 78: 381-384. Langeron M 1949. Précis de Microscopie, 7th ed., Masson & Cie, Paris, 1429 pp. Lunaschi L 1984. Helmintos parásitos de peces de agua dulce de la Argentina. II. Presencia de Lobatostoma jungwirthi Kritscher, 1974 (Trematoda, Aspi-dogastrea) en Cichlasoma facetum (Jenyns). Neotropica 30: 187-192. Oliva ME, Luque JL. 1989. The genus Lobatostoma (Trematoda: Aspidocotylea) in the Pacific coast of South America, with description of Lobatostoma veranoi new species, parasite of Menticirrhus ophicephalus (Teleostei: Sciaenidae). Mem Inst Oswaldo Cruz 84: 167-170. Rai SL 1964. Morphology and life history of Aspidogaster indicum Dayal, 1943 (Trematoda: Aspidobothridae). Indian J Helminthol 16: 100-141. Rohde K 1972. The Aspidogastrea, especially Multicotyle purvisi Dawes, 1941. Adv Parasitol 10: 77-151. Rohde K 1973. Structure and development of Lobatostoma manteri sp. nov. (Trematoda: Aspidogastrea) from the Great Barrier Reef, Australia. Parasitology 66: 63-83. Rohde K 1994. The minor groups of parasitic platyhelminthes. Adv Parasitol 33: 145-234. Rohde K, Sandland R 1973. Host parasite relations in Lobatostoma manteri Rohde (Trematoda: Aspidogastrea). Zeitschrift für Parasitenkunde 42: 115-136. Rohlf FJ, Sokal RR 1981. Biometry. The Principles and Practice of Statistics in Biological Research, 2nd ed., WH Freeman and Company, New York, 859 pp. Szidat L 1971. Weitere Beiträge zur Kenntnis der marinen Reliktfauna des La Plata-Stromsystems. HD Srivastava Commemorative Volume: 637-653. The following images related to this document are available:Photo images[oc99010a.jpg] [oc99010c.jpg] [oc99010d.jpg] [oc99010b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}