 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 94(1): 67-70 RESEARCH NOTE Vaccination of C57BL/10 Mice against Cutaneous Leishmaniasis. Use of Purified gp63 Encapsulated into Niosomes Surfactants Vesicles: A Novel Approach CM LezamaDávila Department of Immunology, University of Strathclyde, Glasgow G4 ONR, U.K. Received 20 February 1998; Accepted 30 September 1998
Code Number:OC99014
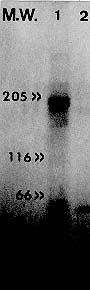
Key words: vaccination - Leishmania m. mexicana - niosomes Leishmaniasis comprises a group of diseases caused by different species of parasites of the genus Leishmania. It is widely accepted that cell mediated immune response plays a major role in the development of this disease, and most experimental evidence indicates that CD4+ T cells are the primary, but not the only population of cells involved in the evolution of this infection in the mouse and, perhaps, in the human being as well (P Scott 1989 Exp Parasitol 68: 369-372, CM Lezama-Dávila et al. 1998 Scand J Immunol 47: 502-508). There is currently no commercial vaccines aimed to prevent L. m. mexicana infections in humans. Although, experimental murine infection induced with Leishmania could be prevented by previous inoculation of attenuated parasites or purified molecules (D Russell et al. 1988 J Immunol 140: 1274-1279, D Rivier et al. 1993 Parasite Immunol 15: 75-81, VF Amaral et al. 1995 Mem Inst Oswaldo Cruz 90 (Suppl. I): 183, CM Lezama-Dávila et al. 1995 In Vivo 6: 519-524, 1997 Arch Med Res 28: 47-53). Moreover, sickness of different strains of mice, excepting C57BL/10 animals, can be effectively prevented by intraperitoneal vaccination with Leishmania surface antigens incorporated into liposomes (Lezama-Dávila et al. 1997 loc. cit.) and this protective effect is transferred by whole Tcells and their subsets (Russell loc. cit., CM Lezama-Dávila et al. 1995 Mem Inst Oswaldo Cruz 90: 51-58). Increase in efficiency of vaccination has been achieved by the use of heterologous carriers (e.g. virus vaccinia or bacteria such as avirulent mutants of Salmonella spp. and BCG) or liposome vesicles that slowly release molecules such as drugs. Niosomes preparations constitute a more recently developed drug delivery system formed by hydration of mono- or di-alkyl surfactants (chemical structure is presented below) that present chemical and physicochemical differences to liposomes, including a better stability and easier handling that makes them attractive as a vesicle-system for antigen release (A Baillie et al. 1985 J Pharm Pharmacol 137: 863-868). This work was designed to test efficiency of vaccination of C57BL/10 mice against cutaneous leishmaniasis by using a novel preparation developed with parasite antigens entrapped into niosomes non-ionic surfactants. We used C57BL/10 mice between 812 weeks old which were bred, sex and age matched in the Department of Immunology of the University of Strathclyde, U. K. Stationary phase promastigotes of L. m. mexicana (MNYC/BZ/62/M379) were obtained by in vitro culture from amastigotes in RPMI1640 supplemented with foetal calf serum, penicillin and streptomycin (Gifco). Mice were infected by subcutaneous inoculation of 3x106 promastigotes into their shaven rumps to induce lesion growth. The day of infection was considered as day 0 of the experiment. Antigens extraction was based on the procedure originally described by Russell (loc. cit.) with slight modifications; 3 X 1010 washed stationary phase promastigotes from culture flasks were disintegrated in a celldisruption chamber. Disrupted parasites were resuspended in 2% octylglucoside in PBS containing 0.05 mM of TLCK, 0.015 mM leupeptide, pH=7.1, maintained in ice for 1520 min and spun down for 1 hr at 100,000 x g in an ultracentrifuge. The supernatant (membrane preparation) was applied to a Con A-Sheparose column. A mannoside gradient was used to elute glycoproteins. Only fractions containing gp63 were detected by ELISA using anti-gp63 MoAbs (hybridoma clones 3.2, 3.8, 3.9; kindly provided by Dr Russell), pooled (gp63) and stored at -20oC. Gp63 containing fractions were analysed by polyacrilamide electrophoresis under reducing conditions using 10% acrilamide to detect molecules with a molecular weight of about 65kD. Antigen incorporation into niosomes was achieved following the procedure of Baillie (loc. cit.). One hundred and fifty mM of single-alkyl chain, non-ionic surfactant (kindly provided by Dr Baillie): C16H33-O-(CH2-CH-O)3-H
|
CH2-OH
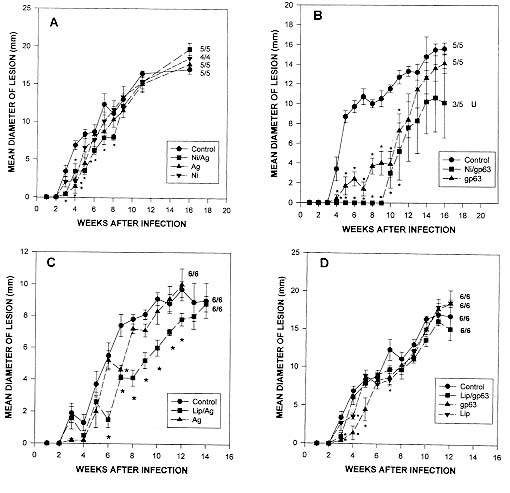
Molecular weight = 473 g/mol and cholesterol in a 100 ml glass container were mixed with 2 ml of antigen (Con A-Sepharose eluted gp63 or membrane antigens) in PBS and sonicated for 3 min. This preparation was washed with PBS and stored at -20oC until use. For the preparation of empty niosomes the same procedure was applied, but PBS without parasite antigen was used. Reconstitution of antigen into liposomes was based on the procedure reported by Russell (loc. cit.): 1 ml suspension 32 mg (80%) soybean lecithin, and 2 mg (5%) diacetyl phosphate were mixed with 2 ml of chloroform in a flask and 6 mg (15%) of cholesterol were added. The mixture was dried with N2 to produce a thin film and incubated for 1 hr at 37oC. Parasite Ag in PBS/octylglucoside was then added to the lipid film and sonicated in an ice bath. Supernatant containing membrane molecules was added and a ratio of 2 mg of protein of parasite sample to 1 mg phospholipid/cholesterol/mixture was maintained throughout the experiments. The detergent was removed by dialysis against PBS at 4oC and the resulting vesicles washed with PBS, centrifuged for 1 hr at 100,000 x g and stored at 20oC until use. The same procedure was applied for the preparation of empty liposomes, except that PBS without parasite antigen was used. Vaccine preparations consisted of membrane molecules or purified gp63 (4 mg of protein/mouse) reconstituted into niosomes or liposomes. These preparations were inoculated twice at two weeks intervals to vaccinate groups of five C57BL/10 mice, which were then challenged (two weeks after the last immunization) with 3 x 106 L. m. mexicana infective promastigotes. Control groups of mice included in this set of experiments were inoculated only with PBS, membrane antigens, purified gp63 or empty niosomes or liposomes before challenge with virulent promastigotes. Our results show that electrophoretic analysis (PAGE-SDS) of membrane antigens presented several strong and weak bands, whereas material eluted from a Con A-Sepharose column and selected by its reactivity with anti-gp63 MoAbs displayed only a strong band with a molecular weight of about 65 kD (Fig. 1). These molecules were encapsulated into niosomes or liposomes and used to vaccinate C57BL/10 mice which were inoculated subcutaneously twice at two weeks interval (4 mg of protein/mouse). Together with unimmunized controls, vaccinated mice were exposed to a homologous challenge (i.e. mice vaccinated with L. m. mexicana gp63 or membrane antigens and infected with the same parasite). Mice subcutaneously vaccinated with membrane antigens reconstituted into niosomes developed lesions at similar rate than control mice. Although, a slight delay (up to week 8 after vaccination) in lesion growth was observed in mice vaccinated with membrane antigens encapsulated into niosomes. The number of animals that developed lesions after 16 weeks of infection was: 5/5 mice subcutaneously vaccinated with membrane antigens reconstituted into niosomes, 5/5 control mice inoculated with empty niosomes, membrane antigens alone or only infected with infective promastigotes (Fig. 2A). There was, however, a considerable disease resistance of C57BL/10 mice when they were vaccinated with purified gp63 reconstituted into niosomes. The number of these mice that developed lesions after 16 weeks of infection was: 5/5 control mice inoculated with purified gp63 alone or only infected with virulent parasites, 3/5 mice vaccinated with purified gp63 reconstituted into niosomes. Additionally, all of these three vaccinated mice presented ulcerated lesions that began to heal. This was in clear contrast to lesions developed by control mice that did not ulcerate or heal by week 16 after infection (Fig. 2B). Fig. 1: polyacrylamide gel electrophoresis of isolated promastigotes antigens following detergent extraction (PAGE-SDS). This figure shows membrane antigens (track 1), Con A-Sepharose purified material of about 65 kD that was selected for its reactivity with different anti-gp63 MoAbs (track 2) and approximate position of molecular weight markers (left), isolated in a silver stained 10% polyacrylamide gel. Fig. 2: effect of intraperitoneal vaccination using membrane antigens (A, C) or Con A-Sepharose purified material of about 65 kD that was selected for its reactivity with different anti-gp63 MoAbs (B, D) reconstituted into niosomes (A, B) or liposomes (C, D) on lesion growth in the shaven rump of C57BL/10 mice. Experimental groups of 5-6 mice were intraperitoneally vaccinated with membrane antigens or gp63 reconstituted into niosomes (squares; A, B) or liposomes (squares; C, D). Control groups (circles) consisted of five mice only infected (sc.) with 3 x 106 promastigotes of Leishmania m. mexicana. Additional controls included the vaccination of mice with empty niosomes (A; inverted triangles), liposomes (D; inverted triangules) or membrane antigens (triangles; A, C) or purified gp63 alone (triangles; B, D). Data are expressed as the mean ± SEM (N=4-6), the number of mice which developed lesions is indicated after each line, ulceration and healing is indicated as needed (U). Significant differences between measurements of control and experimentally vaccinated groups are indicated with asterisks (P<0.01). Data presented represent one of two separate experiments. In addition, groups of six C57BL/10 mice were subcutaneously vaccinated with membrane antigens (Fig. 2C) or purified gp63 (Fig. 2D) alone or reconstituted into liposomes. Mice inoculated with membrane antigens encapsulated into liposomes presented a slight delay in lesion growth between six and 12 weeks after infection. However, 14 weeks after infection all of these animals (6/6) presented lesion diameters similar to controls (Fig. 2C). All animals (6/6) immunized with purified gp63 reconstituted into liposomes developed unhealing lesions at the same rate as control mice (Fig. 2D). Control groups included animals inoculated with empty liposomes or only infected with stationary phase promastigotes (Fig. 2D). Development of a commercial antiparasitic vaccine for human application is an important goal that faces modern Science. We recently published that liposomes improve effectiveness of Leishmania membrane antigens to vaccinate some strains of mice against cutaneous leishmaniasis (Lezama-Dávila et al. 1997, loc. cit.). Nevertheless, this vaccine preparation is not effective to protect C57BL/10 mice against a homologous challenge (Lezama-Dávila 1997, loc. cit.) unless applied together with C. parvum [F Frézard et al. 1995 Mem Inst Oswaldo Cruz 90 (Suppl. I): 194]. Our results on vaccination of C57BL/10 mice indicate that, subcutaneous vaccination against cutaneous leishmaniasis with purified gp63 entrapped into niosomes, but not liposomes, induce considerable resistance to this disease and, at the end of the experiment, these mice presented only ulcerated lesions that started to heal. Control mice, on the other hand, presented only unulcerated lesions with no signs of healing. Recent experiments have also been performed indicating that synthetic peptides deduced from gp63 aminoacid sequence from L. major contains immunoprotective capabilities only when they are encapsulated into pluronic carrier to build up in adjuvanticity (A Jardim et al. 1990 J Exp Med 172: 645-648). Differences in effectiveness of vaccination by using different carriers could result in a different way of presenting Leishmania antigens to the immune system. The latter emphasises once again the importance of a properly selected antigen-release system to develop vaccines. Future research will be necessary to find out immunological pathways followed after vaccination with Leishmania antigens encapsulated into niosomes and possible undesirable side effects in order to assess real possibilities for a vaccination trial in humans. Acknowledgement: to Dr A Baillie for his assistance to develop this work. Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99014a.jpg] [oc99014b.jpg] |
| |||||||||

{kind=link}
{kind=link}