 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 94(1): 71-76 Resolution of an Infection with Leishmania braziliensis Confers Complete Protection to a Subsequent Challenge with Leishmania major in BALB/c Mice Hermenio C Lima/*, Gregory K DeKrey, Richard G Titus/+ Department of Pathology, College of Veterinary Medicine and Biomedical Sciences,
Colorado State University, Fort Collins, CO 80523-1671, USA Received 20 July 1998; Accepted 3 November 1998
Code Number:OC99015
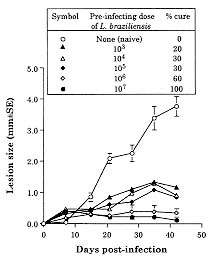
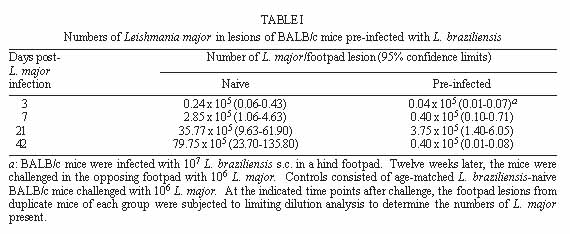
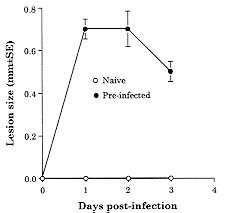
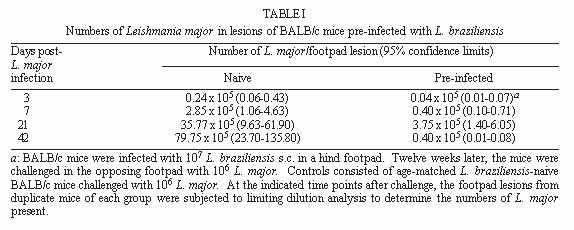
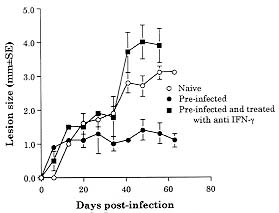
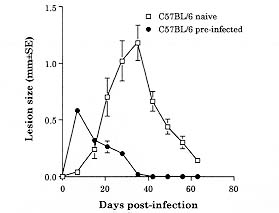
Both Leishmania major and L. braziliensis induce cutaneous leishmaniasis in BALB/c mice. Whereas BALB/c mice die of infection with L. major, they cure an infection with L. braziliensis. We report here that after curing an infection with L. braziliensis, BALB/c mice are resistant to challenge with L. major. When challenged with L. major, L. braziliensis pre-treated BALB/c mice mounted a delayed-type hypersensitivity response to L. major and produced high amounts of interferon-g (IFN-g ) but low amounts of interleukin-4. The IFN-g produced by the L. braziliensis pre-infected mice was involved in the protection seen against L. major challenge since treating the mice with a neutralizing anti-IFN-g abrogated the protection. This suggests that cross-reactive antigen epitopes exist between L. braziliensis and L. major and that pre-infection with L. braziliensis primes BALB/c mice to epitopes on L. major that can elicit a protective Th1 response to the parasite. Key words: Leishmania braziliensis - Leishmania major - mice - cross-protection - cytokines Organisms of the genus Leishmania induce a spectrum of diseases in humans and in experimental animals. Infection of mice with L. major, one cause of cutaneous leishmaniasis, is perhaps the best studied model for cutaneous leishmaniasis (reviewed in Bogdan et al. 1993, Liew & O'Donnell 1993, Reed & Scott 1993, Titus et al. 1994, Reiner & Locksley 1995). Most mouse strains cure an infection with L. major, however BALB/c mice are a notable exception since they ultimately die of infection with L. major when the disease becomes systemic. Considerable work in this model has revealed that mice that are resistant to infection with L. major develop a Th1 immune response and its associated cytokine profile [interferon-gamma (IFN-g)hi; interleukin-4 (IL-4)lo]. IFN-g activates L. major infected macrophages (MØs) to kill the parasite (Murray et al. 1983, Titus et al. 1984, Nacy et al. 1985). In contrast, susceptible BALB/c mice develop a Th2 response and its associated cytokine profile (IFN-glo; IL-4hi). IL-4 can block the ability of IFN-g to activate MØs to kill Leishmania (Lehn et al. 1989, Liew et al. 1989). In contrast to infection with L. major, L. braziliensis induces only a transient cutaneous disease, even in BALB/c mice. This may at least in part be the explanation for why little experimental work has been performed with L. braziliensis (Neal & Hale 1983, Childs et al. 1984). We recently reported (DeKrey et al. 1998) that following infection with L. braziliensis or L. major, BALB/c mice produced similar levels of IFN-g. However, L. braziliensis infected mice produced much less IL-4 (approximately 10-fold). In addition, when the L. braziliensis infected mice were treated with a neutralizing anti-IFN-g, the animals were unable to resolve their infection. We concluded that BALB/c mice cure an infection with L. braziliensis because the low levels of IL-4 they produce are unable to block the ability of IFN-g to activate L. braziliensis infected MØs to kill the parasite. Resolution of an infection with a particular species of Leishmania usually confers complete resistance to re-challenge with the same parasite. However, in addition to this, a primary infection with a given species of Leishmania can also confer cross-protection against a different species of Leishmania (Lainson & Bray 1966, Lainson & Shaw 1977, Alexander & Phillips 1978a,b, Perez et al. 1979, Alexander 1988, Neal et al. 1990, Peters et al. 1990, Melby 1991, Abramson et al. 1995, Gicheru et al. 1997). Cross-protection has been shown in several different mammalian hosts; the protection sometimes acts in only one direction (Lainson & Shaw 1977), and in some cases the sex of the host influences the cross-protection seen (Alexander 1988). Since L. braziliensis is unable to trigger a strong Th2 response in BALB/c mice, we hypothesized that following resolution of an infection with L. braziliensis, BALB/c mice might be at least partially protected against challenge with L. major. We report here that previous exposure to L. braziliensis can confer complete protection against a subsequent challenge with L. major and that this protection is dependent upon IFN-g production. MATERIALS AND METHODS Mice and parasites - Young adult female mice were used in all experiments. BALB/c mice were obtained from either the National Cancer Institute (Bethesda, MD) or Jackson Laboratory (Bar Harbor, ME). C57BL/6 were obtained from the National Cancer Institute. Stationary phase promastigotes of L. braziliensis (MHOM-BR-79-LTB111) or L. major (RHO-SU-59-P) were used. Parasites were maintained as described (Titus et al. 1984). Infecting mice and determining parasite numbers in cutaneous lesions - Mice were injected with the numbers of promastigotes indicated in the text in one hind footpad and lesion development was followed by measuring the thickness of the infected footpad compared to the thickness of the same footpad prior to infection. Parasite numbers were determined in infected footpads using a published limiting dilution assay for determining parasite burdens in infected mouse tissues (Lima et al. 1997). In some experiments mice were treated with a neutralizing anti-IFN-g (XMG1.2) antibody as described in DeKrey et al. (1998). Determining levels of cytokines in culture supernatants - At various times after infection, 3-5 mice per group were killed for evaluation. Single cell suspensions were prepared from draining lymph nodes (inguinal and popliteal). Cells were adjusted to 5x106/ml in Dulbecco's modified Eagle medium (Maryanski et al. 1982) containing 0.5% normal mouse serum (Harlan Bioproducts, Indianapolis, IN). Cultures were stimulated with 106 L. major promastigotes/ml and the supernatant of the cultures was harvested 72 hr later (a time determined to be optimal for the cytokines examined) for analysis. Levels of IFN-g and IL-4 in culture supernatants were determined by enzyme-linked immunosorbent assay (ELISA) using techniques published elsewhere (Soares et al. 1997). Statistical analysis - Significance was determined using an non-paired t test. Differences were considered to be significant when p < 0.05. All experiments shown are representative of two to three independent experiments. RESULTS To determine whether previous exposure to L. braziliensis led to protection against a subsequent challenge with L. major, we first experimented with the dose of L. braziliensis and the time between infection with L. braziliensis and challenge with L. major. We found that a large dose of L. braziliensis (107) administered subcutaneously (s.c.) in one hind footpad led to complete protection against a subsequent challenge with 106 L. major s.c. in the opposing hind footpad (Fig. 1). Moreover, the protective effect of pre-infecting with L. braziliensis was a dose titratable phenomenon. As shown in Fig. 1, a dose of 103 L. braziliensis led to the least protection against challenge with L. major whereas a dose of 107 L. braziliensis led to the greatest protection. Lesions of L. major were the largest in mice pre-treated with 103 L. braziliensis and only 20% of the mice (see numbers in the legend of Fig. 1) cured these L. major-induced lesions; in contrast, lesions of L. major were the smallest in mice pre-treated with 107 L. braziliensis and 100% of the mice cured these L. major-induced lesions. Fig. 1: course of infection with Leishmania major in BALB/c mice pre-infected with different concentrations of L. braziliensis. Groups of 10 BALB/c mice each were pre-infected with the indicated doses of L. braziliensis s.c. in one hind footpad. Twelve weeks later these animals were challenged s.c. in the opposing hind footpad with 106 L. major. Controls consisted of naive mice infected with 106 L. major. Lesions were monitored as described in Materials and Methods. We also determined that the degree of resistance to challenge with L. major increased with time after exposure to L. braziliensis. Little if any protection against challenge with L. major was achieved when the two parasites were injected simultaneously. Some protection was observed when mice were challenged with L. major at 6 or 8 weeks after exposure to L. braziliensis. However, 100% protection against challenge with L. major was consistently achieved only at 12 weeks after exposure to L. braziliensis (data not shown). Importantly, at 12 weeks post-L. braziliensis injection, we were also unable to detect viable L. braziliensis in treated mice by limiting dilution analysis (data not shown). Therefore, for the remaining experiments presented here, mice were treated with 107 L. braziliensis and challenged 12 weeks later with 106 L. major. The experiment shown in Fig. 1 demonstrated that pre-infection with L. braziliensis allows BALB/c mice to control the outgrowth of lesions of L. major when the mice were challenged with the parasite. To determine whether this was accompanied by destruction of L. major in the lesions, we measured the parasite burdens in the lesions. In L. braziliensis-naive control mice, L. major continued to replicate through day 42 of infection (Table I). In contrast, in mice pre-infected with L. braziliensis 12 weeks earlier, L. major was destroyed such that by day 42 of the experiment there were approximately 2,000-fold fewer parasites in their lesions compared to control mice (Table I). We next analyzed the mechanism underlying the protection seen against challenge with L. major in BALB/c mice pre-infected with L. braziliensis. We first noted that an intense swelling response occurred in the footpads of L. braziliensis pre-treated mice when the mice were challenged with L. major (Fig. 2). This swelling response was characteristic of delayed-type hypersensitivity (DTH) in that it peaked from 24 to 48 hr post-challenge with L. major and it persisted to 72 hr post-challenge (Fig. 2). This observation suggested that cross reactive antigenic epitopes exist in L. braziliensis and L. major that prime T cell responses. Moreover, since DTH is mediated by Th1-type T cells (Mosmann & Coffman 1989), this also suggested that infection with L. braziliensis triggered Th1 T cells in BALB/c mice that could recognize L. major antigen(s) when the mice were challenged with the parasite. Fig. 2: footpad swelling response of Leishmania braziliensis pre-infected BALB/c mice challenged with L. major. BALB/c mice were pre-infected with L. braziliensis and challenged with L. major as described in the legend of Fig. 1. To test the hypothesis that cross reactive Th1 T cells were elicited by pre-infection with L. braziliensis, we measured the cytokines produced when lymph node cells from L. braziliensis pre-infected mice were challenged with L. major in vitro. We first harvested the popliteal and inguinal nodes draining the footpad of mice pre-infected with L. braziliensis 12 weeks earlier. These cells were stimulated with L. major promastigotes in vitro and the supernatants were harvested 72 hr later to determine their content of IFN-g and IL-4. These lymph node cells produced substantial amounts of IFN-g (15.48 ng/ml, Table II) but no detectable IL-4. Moreover, when the same lymph node cells were harvested from L. braziliensis pre-infected mice at varying times after the mice were challenged with L. major, the cells continued to produce substantially more IFN-g but less IL-4 than control naive BALB/c mice challenged with L. major (Table II). Since L. braziliensis pre-infected mice produced elevated levels of IFN-g but lower levels of IL-4 compared to naive mice when the mice were challenged with L. major (Table II), we next tested the hypothesis that the IFN-g was involved in the protection seen against challenge with L. major. L. braziliensis pre-infected BALB/c mice were treated with a neutralizing IFN-g antibody as described in the Materials and Methods and challenged with L. major. As can be seen in Fig. 3, treating with anti-IFN-g abrogated the protection seen when the mice were challenged with L. major. Fig. 3: Leishmania braziliensis pre-infected BALB/c mice do not resist infection with L. major when the animals are treated with anti-IFN-g antibody. Groups of BALB/c mice were pre-treated with 107 L. braziliensis and 12 weeks later the mice were challenged with 106 L. major. One of the groups of pre-treated mice was also injected with a neutralizing anti-IFN-g antibody as described in Materials and Methods. Controls consisted of age-matched L. braziliensis-naive BALB/c mice challenged with L. major. Lesions were monitored as described in Materials and Methods. Finally, since L. braziliensis pre-infected BALB/c mice were protected against challenge with L. major, we tested whether resistance to infection with L. major was enhanced in a mouse that normally cures an infection with the parasite. C57BL/6 mice were pre-infected with 107 L. braziliensis and 12 weeks later were challenged with 106 L. major. As can be seen in Fig. 4, these mice showed markedly increased resistance to challenge with L. major. Fig. 4: pre-infecting with Leishmania braziliensis enhances the resistance of C57BL/6 mice to challenge with L. major. C57BL/6 mice were pre-infected with 107 L. braziliensis promastigotes and 12 weeks later the mice were challenged with 106 L. major. Controls were L. braziliensis-naive C57BL/6 mice challenged with L. major. Lesions were monitored as described in Materials and Methods. DISCUSSION We recently reported that cutaneous lesions develop on BALB/c mice following infection with either L. major or L. braziliensis; however, while BALB/c mice ultimately die of infection with L. major, the mice resist infection with L. braziliensis, kill the parasite, and heal their cutaneous lesions (DeKrey et al. 1998). Further analysis of this system revealed that following infection with L. major, BALB/c mice developed a Th2-biased immune response. In contrast, following infection with L. braziliensis, the mice developed a Th1-biased response (DeKrey et al. 1998). Since a Th1 response mediates cure in mice infected with L. major, we hypothesized that BALB/c mice that had cured an infection with L. braziliensis would resist challenge with L. major. To our knowledge, no one has examined whether pre-infecting mice with L. braziliensis confers protection against challenge with L. major. We report here that pre-infecting BALB/c mice with L. braziliensis confers full protection against challenge with L. major. That is, whereas L. braziliensis-naive control mice were susceptible to infection with L. major, L. braziliensis pre-treated mice resolved cutaneous lesions of L. major (Fig. 1) and destroyed L. major parasites within the lesions (Table I). Full protection against L. major required pre-infection with a high dose of L. braziliensis (107, Fig. 1). The reason such a high dose of L. braziliensis was required is not known. However, L. braziliensis is known to be poorly infective for laboratory mice (Samuelson et al. 1991). Since the metacyclic form of Leishmania is the infective form of the parasite [the form that survives and efficiently infects MØs in the vertebrate host (da Silva et al. 1989, Puentes et al. 1990)], it is possible that conversion to metacyclics by L. braziliensis is very inefficient using standard culture conditions. Thus, a large dose of L. braziliensis is required to successfully infect antigen presenting cells such as MØs in the host which in turn stimulate an effective immune response. In addition to requiring a large dose of L. braziliensis to achieve full protection against challenge with L. major, it was also necessary to wait 12 weeks before challenging with the parasite, a time when L. braziliensis parasites could not be detected in the mice. Indeed, little if any protection was achieved when L. braziliensis and L. major parasites were injected simultaneously. Taking these observations together, in conjunction with the fact that L. braziliensis pre-treated mice produced large amounts of IFN-g when challenged with L. major (Table II), suggests that protection against challenge with L. major was not mediated by either concomitant immunity or a non-specific inflammatory response against L. major. Rather, the data suggest that there are antigenic epitopes shared between L. braziliensis and L. major. When a BALB/c mouse is infected with L. braziliensis, these epitopes elicit a protective Th1 T cell response such that the mice mount a Th1 response when challenged with L. major. The nature of the cross-reactive epitopes of L. braziliensis and L. major are unknown but are currently under investigation. It would be interesting if these cross-reactive epitopes were found to be expressed at low levels on L. braziliensis and/or to stimulate rare T cell clones. If this were the case, it would offer an alternative explanation for our observations that a large dose of L. braziliensis was required to achieve full-protection against L. major challenge and that it required 12 weeks for this protection to develop. In conclusion, the data presented here confirm and further characterize previous reports that demonstrated cross-protection between different species of Leishmania. In addition, the data show that BALB/c mice can be induced to mount a protective Th1 response against a normally lethal infection with L. major, and that this can occur in the absence of intervention with cytokines or anti-cytokines. ACKNOWLEDGMENTS To Monica Estay for excellent technical assistance. This work was supported by the National Institutes of Health grant AI 29955. REFERENCES
Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99015b.jpg] [oc99015e.jpg] [oc99015d.jpg] [oc99015c.jpg] [oc99015a.jpg] [oc99015f.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}