 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Influence of Growth Medium in Proteinase and Pigment Production by Streptomyces cyaneus Simone DG Petinate, Rosana M Martins, Rosalie RR Coelho, Maria Nazareth L Meirelles*, Marta Helena Branquinha, Alane Beatriz Vermelho/+ Departamento de Microbiologia Geral, Instituto de Microbiologia
Professor Paulo de Góes, Universidade Federal do Rio de
Janeiro, CCS, Bl. I, 21949-900 Rio de Janeiro, RJ, Brasil + Corresponding author. Fax: +55-21-560.8344. E-mail: immgbia@microbio.ufrj.brThis work was supported by CNPq, FUJB, Finep-BID, CEPG-UFRJ, Fiocruz. Received 2 April 1998 Code Number:OC99033 RESEARCH NOTE Key words: Streptomyces - proteinase - melanoid pigment - catabolite repression Several species from the Streptomyces genus produce bioactive molecules, including enzymatic inhibitors with antibiotic activity, and many commercial valuable enzymes such as lipases, cellulases and proteases (W Piepersberg 1994 Crit Rev Biotech 14: 251-285, W Peczynska-Czoch & M Mordarski 1988, p. 219-283. In M Goodfellow, ST Williams, M Mordarski (eds), Actinomycetes in Biotechnology, Academic Press, London). This last class of proteins has been found to have application in the pharmaceutic, detergent and food industries as well as in laboratory practice (TP Lyons 1988 Crit Rev Biotech 8: 99-113, CA Etok & OU Eka 1996 J Basic Microbiol 36: 83-88, P Clapés et al. 1997 Biotechnol Lett 19: 1023-1026). Many of the extracellular proteinases obtained from streptomycetes have been characterized as metallo- and serine-proteinases (K Kitadokoro et al. 1994 Eur J Biochem 220: 55-61, SG Kang et al. 1995 Microbiol 141: 3095-3103, IS Kim & KJ Lee 1996 Microbiol 142: 1797-1806). It appears that they are involved in the nutrition, degradation of aerial mycelia and sporulation processes. In addition, this group of actinomycetes is also able to synthesize and excrete dark pigments, the melanins or melanoid pigments, which are considered as an useful criterium in taxonomic studies (T Arai & Y Mikami Y 1972 Appl Microbiol 23: 402-406, ST Williams et al. 1983 J Gen Microbiol 129: 1743-1813). Therefore, the characterization of components secreted by actinomycetes may be useful in many biotechnological fields. In the present study, we have evaluated the effect of different
culture media and substrates on the Streptomyces cyaneus
proteinase and pigment production. This microrganism was isolated
from Brazilian cerrado soil, a particular kind of soil whose major
features are the organic matter-poor content and acidic pH (5.5),
and was maintained in glycerol-peptone-salt liquid medium. The
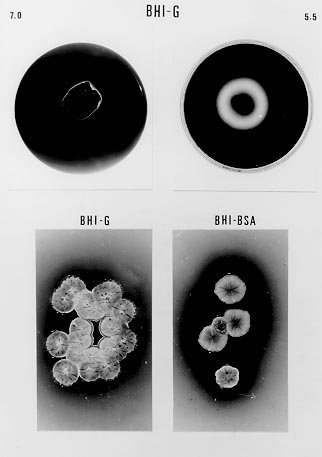
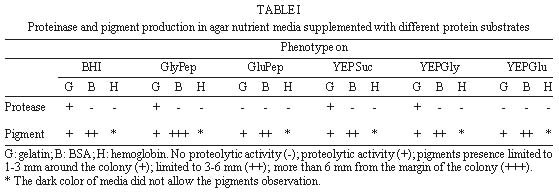
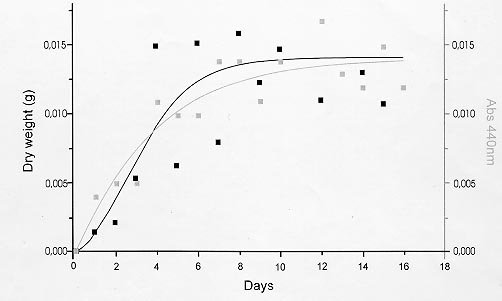
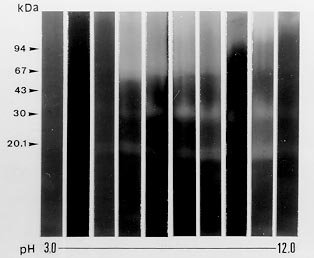
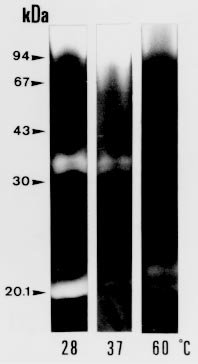
following media containing 1.5% agar were used: In order to detect extracellular proteinase and pigment production on nutrient-agar plates, the six media were supplemented with 1% of the following proteins: bovine serum albumin (BSA), hemoglobin and gelatin. Twenty ml of each culture medium were then poured per plate and allowed to harden (AB Vermelho et al. 1996 Mem Inst Oswaldo Cruz 97: 755-760). A loopful of the culture growth in glycerol-peptone-salt medium was placed in the center of the agar plates. After inoculation, the plates were incubated at 37ºC and observed daily for ten days. To detect the presence of proteinases, the plates were stained with 0.1% amido black in methanol-acetic acid-distilled water (30:10:60, vol/vol/vol) and destained with water until the appearance of clear zones around growth. All the experiments were performed in triplicate and the influence of pH (5.5, 7 and 10) on the proteinase synthesis was evaluated using the BHI and YEPSuc media supplemented with gelatin 1%. The pH of both media was adjusted before steam sterilization with 0.2 M sodium phosphate and 0.4 M glycine/0.1 M NaOH buffer solutions. The formation of dark-brown diffusible pigments and culture growth characteristics on the various solid culture media were macroscopically observed. Additionally, S. cyaneus extracellular proteolytic activity was analyzed in GlyPep liquid medium. For proteinase production and growth studies, 4 ml from a suspension containing 105 spores/ml was added to 200 ml of the liquid broth and incubated for 16 days at room temperature. Ten ml daily aliquots of culture medium were collected and centrifuged at 1500 g for 30 min. Cells were washed with 0.85% NaCl solution and liophylized for biomass determination. Extracellular proteolytic activity was assayed using azocasein as substrate (G Sarath et al. 1989, p. 22-55. In RJ Beynon & JS Bond (eds), Proteolytic Enzymes - A Practical Approach, IRL Press, Oxford). A reaction mixture containing 1 ml of culture supernatant, 1 ml of Tris-HCl 0.1 M, pH 9, 200 m l of CaCl2 0.2 M and 50 m l of azocasein (0.1 mg/ml) was incubated at 37oC for 1 hr. The reaction was stopped by the addition of trichloroacetic acid 5% (1.5 ml) and the proteolytic activity defined as the increase of one absorbance unit at 440 nm. Effect of temperature and pH on extracellular proteolytic activity was evaluated by SDS-PAGE 10% containing gelatin (1%) as substrate (C Heussen & EB Dowdle 1980 Anal Biochem 102: 196-202). Nine-day cell-free culture supernatant was concentrated against polyethyleneglycol 4000 at 4oC overnight and mixed with sample buffer (UK Laemmli 1970 Nature 227: 680-685) in a proportion of 8:2 (vol/vol). The temperature effect was assayed by incubating gel strips for 24 hr at 0.4 M glycine-0.1 M NaOH, pH 10, at 28oC, 37oC and 60oC. The pH effect on proteolytic activity was evaluated at 28oC using the following buffers: 0.1 M citrate (pH 3-5), 0.1 M phosphate (pH 6-8), 0.1 M Tris-HCl (pH 9) and 0.4 M glycine-0.1 M NaOH (pH 10-12). In order to investigate the class of the extracellular proteinases, concentrated cell-free culture supernatant was incubated with proteolytic inhibitors at room temperature for 1 hr prior to addition of azocasein. Concentrations of the inhibitors during the pre-incubation step were indicated in the Table II. Table I shows the effects of different culture media and proteinaceous subtrates addition on S. cyaneus proteinase and pigment production. Proteases halo were observed in all plates containing gelatin as substrate, except for glucose-containing media. Hemoglobin and BSA were not hydrolysed by the extracellular proteinases. However, a clear region under the colonies was observed when BSA was used (data not shown). The fact that S. cyaneus extracellular proteinases preferentially used gelatin, but not hemoglobin and BSA, may suggest the presence of specific cleavage sites in this protein. When glucose was added to the culture media, no proteolysis halo was detected. Thus, it appears that this carbohydrate induces an inhibitory effect on proteinase synthesis similar to that observed in other Streptomyces sp. (IS Kim & KJ Lee 1995 J Biotechnol 42: 35-44, Kim & Lee 1996 loc. cit.). In these and other microrganisms the phenomenon which influences the biosynthesis of many microbial products in solid-state cultivation has been described as catabolite repression (I Delic et al. 1992 Proc Ntl Acad Sci USA 89: 1885-1889, L Servin-González et al. 1994 Microbiol 140: 2555-2565, JM Gancedo 1998 Microbiol Mol Biol Ver 62: 334-361). However, whether the prevention of protease secretion observed in this study in liquid culture is in fact a phenomenom of catabolite repression remains to be demonstrated. Although the BSA was not hydrolysed by the extracellular proteinases of S. cyaneus, this protein was an excellent substrate for the production of melanoid pigment. Moreover, BHI-BSA and YepGly-BSA induced a weaker pigmentation, but in GlyPep-BSA the pigmentation was stronger (Fig.1, Table I). Melanin biosynthesis in actinomycetes may occur either in a proteic medium, specifically, in a tyrosine-containing medium, or in a synthetic medium, without tyrosine or protein addition, but instead with an inorganic N source. When the melanin synthesis occurs in a proteic medium, the phenoloxidase enzyme, tyrosinase, transforms L-tyrosine into L-DOPA, and this into dopachrome, which in turn is auto-oxidized to indol-5,6-quinone. The later is polymerized spontaneously into DOPA-melanin. In the second case, the pigments are produced from the metabolic products of the microrganisms, such as aromatic compounds or quinones, or even from antibiotics with quinone structures, produced during secondary metabolism (GA Swan 1974 Fortschr Chem Org Naturst 31: 521-582, RRR Coelho & LF Linhares 1993 Biol Fert Soils 15: 220-224, RC Gomes et al. 1996 Biol Fert Soils 21: 84-88). In our case the increased pigment production induced by the presence of BSA may be associated to the transformation of aromatic amino acids, which are present in high amount in the structure of this protein. FIGURE 1: influence of brain heart infusion (BHI) medium, pH and proteic substrates on proteinases and melanoid pigment production secreted by Streptomyces cuaneus growth in solid state medium. pH values are indicated in the left and right upper sides of the figure. G: gelatin, BSA: bovine serum albumin. TABLE I: Proteinase and pigment production in agar nutrient media supplemented with different protein substrates. The pH of the culture medium has a marked effect on the protease production during the microrganism growth. BHI-gelatin at pH 5.5 was the best medium for proteolytic activity detection on agar plates (Fig. 1). Taking into consideration that Brazilian cerrado soil is an acidic tropical soil, the secretion of proteolytic enzymes in acidic pH by S. cyaneus is an important feature in the degradation of vegetal and animal proteins and, consequently, in humus formation. In addition, it has been observed that extracellular protease production is more intense in streptomycetes isolated from Mata Atlântica, an organic matter-rich soil, than from cerrado soil (unpublished data). Probably the small amount of extracellular proteinases in this microrganism is due to the low level of organic matter in cerrado soil, exerting a highly selective pressure in this environment. Similarly, D Jendrossek et al. (1997 FEMS Microbiol Lett 150: 179-188) demonstrated that soil samples of the rubber-tree Hevea brasiliensis plantations and waste ponds of rubber-producing companies are rich in natural rubber-degrading bacteria. Morphological alterations in the colonies were induced by the added substrates and pH of the media. Hemoglobin addition in nutrient-agar led to the formation of mucoid colonies, and alkaline media decreased the size of the colonies, forming very small ones (data not shown). S. cyaneus proteinase production and growth studies showed a significant increase of proteolytic activity from the second day of culture. The proteolytic activity of the medium increased in parallel to the bacterial growth until the culture reached the stationary phase (Fig 2). These results are in accordance with previous observation in other Streptomyces species (Kim & Lee 1996 loc. cit., Kang et al. 1995 loc. cit.) In addition, a serine proteinase appearing in the earling logarithmic phase of growth was associated to the morphological diferentiation of these microrganisms. FIGURE 2: secretion of proteinases during Streptomyces cyaneus growth in GlyPep liquid-medium. Growth was measured as dry weight and proteolytic activity assayed using azocasein as substrate. Aliquots were collected from the first to the sixteenth day of culture. When the supernatant was ten times concentrated and analyzed by SDS-PAGE-gelatin two bands (30 kDa and 20 kDa) of activity could be observed (Figs 3, 4). The molecular weight of these proteinases bands are identical to that found to other enzymes from Streptomyces genus (KH Yeoman & C Edwards 1997 J Appl Microbiol 82: 149-156). In addition, a smear of proteolysis activity in the top of the gel was seen (Figs 3, 4). Since it is known that many environmental factors such as temperature and pH may affect the aggregation state of secreted proteins, it is possible that this high molecular weight activity may be due to a multimeric form of the proteinase. FIGURE 3: effect of the pH on Streptomyces cyaneus extracellular proteolytic activities anlyzed by SDS-PAGE-gelatin. Gel strips were incubated for 24 hour at different pH values (3-12). Molecular weight markers in kDa are indicated in the left side of the figure. FIGURE 4: effect of the temperature on Streptomyces cyaneus extracellular proteolytic activities detected by SDS-PAGE-gelatin. Gel strips were incubated for 24 hr at different temperature (28° , 37° , 60° C) and the staining developed s described early. Molecular weight markers in kDa run simultaneously in the gels and are indicated in the figure. The activity of the proteinases was more pronounced at pH 9-11 and in a temperature of 28oC (Figs 3, 4). The 20 kDa protein was resistant for 1 hr at 60oC in the conditions used in this study. Almost no activity was detected at pH values below 6 and at pH 12. In this study, two distinct culture conditions of growth were used. S. cyaneus was cultured either in BHI-agar at ph 5.5 or in GlyPep liquid medium at pH 7. The pH at which the culture was grown influenced the activity that was found. The acidic proteinases induced at pH 5.5 are probably different from the proteolytic activities detected at pH 7. This observation has already been described for yeast extracellular proteases, in which the pH of culture medium influenced the optima pH conditions for enzymatic activity (DM Ogrydziak 1993 Crit Rev.Biotechnol 13: 1-55). Extracellular proteolytic activity was partially inhibited by PMSF and STI, while EDTA, EGTA, E-64 and pepstatin had no effect (Table II). These results indicate that both enzymatic activities detected in S. cyaneus may belong to the serine-proteinases group. TABLE II: Effect of various proteolytic inhibitors on extracellular proteinases activity measured using azocasein as substrate. The results represents the men of three experiments Finally, this work reports our earlier results on proteinases secreted by a streptomycete isolated from Brazilian cerrado soil. Temperature-resistant and some alcalophylic properties of Streptomyces sp. proteases are of sum importance in several industrial and biotechnological applications. Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99033c.jpg] [oc99033d.jpg] [oc99033a.jpg] [oc99033b.jpg] [oc99033f.jpg] [oc99033e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}