 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Wild Birds as Hosts of Amblyomma cajennense (Fabricius, 1787) (Acari: Ixodidae) Rosario Rojas/+, Miguel Ângelo Marini*, Maria Teresa Zanatta Coutinho** Curso de Pós-graduação em Ecologia,
Conservação e Manejo de Vida Silvestre
+ Received 24 July 1998
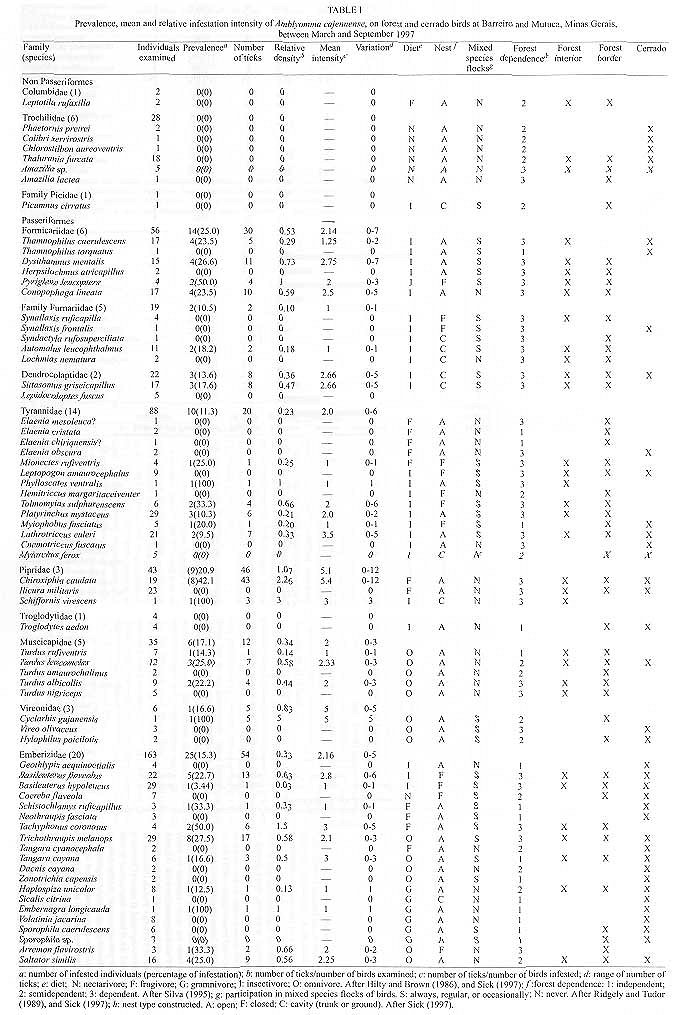
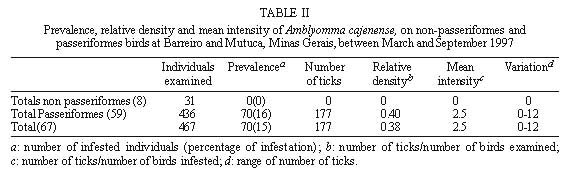
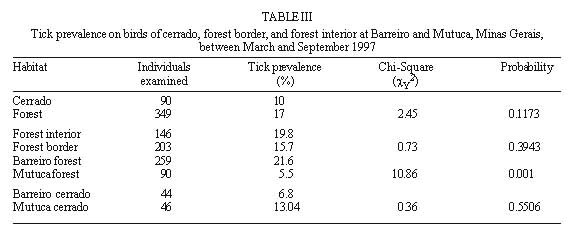
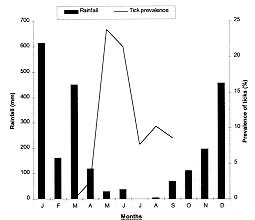
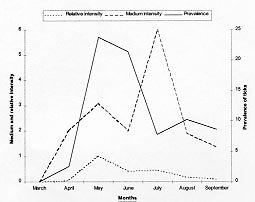
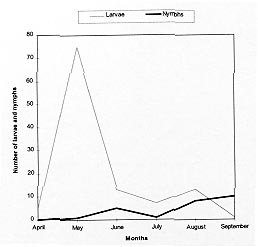
Code Number:OC99060 We evaluated the prevalence, mean intensity and relative density of ticks in 467 wild birds of 67 species (12 families) from forest and cerrado habitats at two protected areas of Minas Gerais, between March and September 1997. Ticks collected (n=177) were identified as larvae and nymphs of Amblyomma cajennense and four other species of Amblyomma. We report for the first time 28 bird species as hosts of the immature stages of A. cajennense, demonstrating the lack of host specificity of the larvae and nymphs. A. cajennense had 15% prevalence on birds, with a mean infestation intensity of 0.37 ticks per host sampled, and 2.5 ticks per infested bird. Prevalence varied in relation to host species, diet and between birds from forests at two successional stages. There were no differences in relation to host forest dependence, participation in mixed flocks of birds, and nest type constructed. A. cajennense is a species of medical and veterinary importance, occurring on domestic animals but is known little of its occurrence on wildlife. Key words: Amblyomma cajennense - birds - wild hosts - larvae - nymphs Amblyomma cajennense (Fabricius, 1787) is an ixodid tick which parasitizes man, domestic and wild animals and is very common in Brazil. It occurs in the New World from southern South America to southern North America (Hoogstraal & Aeschlimann 1982) and is a known vector of Rickettsia rickettsii which causes Rocky mountain spotted fever in humans as well as being a possible vector of brucella and Trypanosoma cruzi in humans (Smith 1974). It is also a possible vector of piroplasmosis in cattle and has been demonstrated to cause hematological alterations in parasitized cattle (Serra Freire & Cunha 1987). The life cycle of A. cajennense has four stages: egg, hexapod larvae, octopod nymph and adult (Walker 1994). The cycle of A. cajennense during its free stages and as a parasite of domestic species has been studied primarily under controlled laboratory conditions by Rohr (1909), Olivieri and Serra Freire (1984a, b), Souza and Serra Freire (1992), Serra Freire and Olivieri (1992), and Lopes et al. (1998) A. cajennense is considered an eclectic ectoparasite, both during its immature stages and possibly during its adult stages, with a wide range of hosts (Hoogstraal & Aeschlimann 1982). Adults usually parasitize equids, cattle and dogs as well as several other domestic species. This species has also been found on several wild vertebrates (Aragão 1936), including rodents (Linardi et al. 1987), edentates (Botelho et al. 1989) and several other species of small mammals (Coutinho 1997). There is lack of studies on the role of wild birds on the dispersal of immature stages of A. cajennense. This study presents data on wild birds as hosts and dispersers of immature stages of A. cajennense. It evaluates the prevalence, mean intensity and relative density of infestation on birds from forests and cerrado habitats of Minas Gerais, correlating the infestation indexes with host taxonomy, environmental variables and ecological characteristics of the birds. MATERIALS AND METHODS This study was conducted at "Áreas de Proteção Especial (APE) para fins de Preservação de Mananciais" owned by "Companhia de Saneamento de Minas Gerais" (Copasa), (43o.50'W, 19o.50'S). Two study areas were sampled (1) Mutuca APE (1,250 ha) and (2) Barreiro APE (1,406 ha), located in the municipalities of Nova Lima and Belo Horizonte, respectively. Vegetation at these areas is composed of cerrado, gallery forest, and dry forest (Cetec 1996). At Barreiro APE, the forest is a more developed dry tropical forest at a more advanced successional stage (~ 150 yrs) than the Mutuca APE forest which has gallery and dry forest at an earlier successional stage (~ 90 yrs) (Cetec 1996). The region has distinct seasonal weather changes with warm rainy summers between November and March and cool dry winters between April and October (Cetec 1983) (Fig. 1). During 1997, the study sites had a total precipitation of 2,244 mm. The mean air temperature varied seasonally between 18oC and 24oC, July being the coolest month with mean air temperature between 14oC and 22oC (Sistema de Controle Climatológico Copasa). Birds were captured at forest fragments of 1, 2, 32, and 150 ha at Barreiro and at a 200 ha fragment at Mutuca. Areas sampled included forest interior, the edge of these forests and cerrado transects in Mutuca and Barreiro. Captures were conducted between March and September 1997, with 15-17 mist nets (36 mm mesh, and 12 m long by 2,5 m high), placed close to the ground and opened between 7:00 and 16:00 three to four days a week, in a different area each week. Birds were identified with the help of field guides (Hilty & Brown 1986, Dunning 1987, Ridgely & Tudor 1989 a, b). Each bird captured was weighed, metal and color banded, measured (tail, wing, tarsus, and culmen), checked for the presence of molt, brood patch, sexed when possible, and aged through cranial ossification. Ticks were sampled checking mostly the head, neck, and belly, because these are the areas were ticks are most often found attached to birds. Ticks were collected from the body of birds with forceps and preserved in 70% alcohol. Birds were released after sampling. For the identification of ticks we used the Clifford and Anastos (1960) and Famadas et al. (1997). Each bird species had its prevalence (number of individuals of a host species infected with a parasite species divided by the number of hosts examined), and its mean intensity (mean number of individuals of a parasite species per infected host in a sample) and relative density (mean number of individuals of a parasite species per host examined) (Margolis et al. 1982). Prevalence was compared with respect to bird taxon, diet, participation in mixed species flocks (feeding aggregations of birds), nest type constructed, and sex. Environmental variables such as level of forest dependence, forest edge effects, forest size effects, and seasonal variation, were also tested for differences in tick prevalence. The prevalence of ticks on birds and the variables considered were tested with Chi-square tests using Yates correction (c Y2) when degrees of freedom were 1. Spearman rank correlation (rs) was used to compare prevalence and forest size and prevalence and medium infestation intensity. Statistical analyses were considered significant at 5%. RESULTS Tick prevalence, mean intensity and relative density were evaluated for 436 birds of 59 species (9 families) of Passeriformes and 31 bird species (3 families) of non-Passeriformes (Tables I, II). Ticks collected (n=177) were identified as larvae and nymphs of A. cajennense and also of four other unidentified species of Amblyomma. Adults were never found on birds. A. cajennense was present on 28 bird species ( Table I), and four other unidentified Amblyomma species were found on the following birds: Turdus albicollis, Pyriglena leucoptera, Tolmomyias sulphurescens, Basileuterus flaveolus, Cono-pophaga lineata and Saltator similis. Passeriformes and non-Passeriformes showed differences in prevalence values, since no ticks were found on the 31 non-passerine birds sampled whereas the passerines had a prevalence of 16% ( Table II). Prevalence values varied among bird families, being highest on Formicariidae (25%) and Pipridae (20.9%). The values of the mean infestation intensity were high for all passeriform families, being highest on Pipridae (5.1), Vireonidae (5.0) and Dendrocolaptidae (2.66). The highest relative infestation intensity was detected on Pipridae (1.07), Vireonidae (0.83) and Formi-cariidae (0.53) (Table I). Tick prevalence varied significantly (c 2=64.573; gl=4; p=0.000) among bird dietary groups being highest on omnivorous (21.2%) and lowest on nectarivorous (0%) birds. Birds that participate in mixed-species flocks did not differ in tick prevalence (c Y2=0.16; df=1; p=0.691) from those that do not participate. Also, birds nest type did not show any significant relationship (c 2=0.449; df=2; p=0.975) with A. cajennense prevalence. Tick prevalence also did not vary in relation to host sex. This pattern was evident both for the species with more than 10 individuals sampled as well as for males and females of all species pooled together (c Y2=0.38; df=1, p=0.539). Birds with different levels of forest dependence did not show different prevalence of A. cajennense (c 2 =2.73; df=2; p=0.200). No significant difference was detected in tick prevalence between birds captured at the interior and at the edge of the forests ( Table III). Tick prevalence also did not vary significantly between birds captured at forests and at cerrado formations ( Table III). The correlation between tick prevalence and forest size was also not significant (rs= 0.60; p=0.284). The birds from the older forest (Barreiro) had higher mean tick numbers than the birds at the younger forest (Mutuca). The highest prevalence values were observed at the months with the lowest precipitation (Fig. 1). The infestation peak ocurred in May and declined in March and July. The mean infestation intensity was highest in July, when prevalence was lowest, showing that infestation was intense on the few individuals infested (Fig. 2). Nymphs were uncommon during the months sampled with the highest values in August and September, whereas the larvae were more abundant than the nymphs and had its highest ocurrence in May (Fig. 3). There was no significant correlation between prevalence and mean infestation intensity neither at the family (n=8, r=0,479 p=0.229) nor at the species (n=28, r=0.181 p=0.354) level. Fig. 1: rainfall and Amblyomma cajennense prevalence on birds at Barreiro and Mutuca, Minas Gerais during 1997 Fig. 2: Amblyomma cajennense prevalence and medium and relative intensity on birds at Barreiro and Mutuca, Minas Gerais, between March and September 1997. Fig. 3: number of larvae and nymphs of Amblyomma cajennense on birds at Barreiro and Mutuca, Minas Gerais, between April and September 1997. DISCUSSION The birds sampled seem to be secondary hosts helping in the dispersal of immature phases (larvae and nymphs) but not adults of A. cajennense. A. cajennense occurred on 28 of the 67 species of birds sampled, showing a lack of host specificity during its immature stages, as suggested by Hoogstraal and Aeschlimann (1982). The hosts of the adult phase of A . cajennense include several species of wild and domestic vertebrates (Aragão 1936). Prevalence of Amblyomma was of 15% for all birds sampled, with 0% prevalence for non-Passeriformes and 16% for Passeriformes(Table I). These values are similar to the ones reported by Pruett-Jones and Pruett-Jones (1991) for other species of Ixodid in forest birds of New Guinea (15.4% for all birds; 0.6% for non-Passeriformes; and 16.4% for Passeriformes). However, the total values differ from the ones reported by Marini et al. (1996) for Passeriformes from the Atlantic Forest of the State of Paraná (45.5%) and from Marini and Couto (1997) for Passeriformes and non-Passeriformes captured at several forests of Minas Gerais (24.3%). These data reveal a wide variation in the prevalence of ixodid ticks on birds among different geographical regions. Our analysis also showed that there is variation in the infestation level of Amblyomma, depending on the ecology and behavior of the host. The pattern of infestation ranges from host species with high infestation levels to species which are uninfected. This is evident for Passeriformes which had high mean infestation levels and especially in some families (Table I). Host species with the highest prevalence also showed high mean infestation intensity at both the family and species level. This pattern was also reported by Pruett-Jones and Pruett-Jones (1991) and Marini et al. (1996). High infestation may represent low immunity in some individuals, a decrease in health state and body condition, and a decrease in the birds' movement ability which may increase the encounter rate with ticks, causing a further increase in the infestation probability (Pruett-Jones & Pruett-Jones 1991). The low prevalence of A. cajennense at the sampled birds may be related to their low host specificity and to the fact that A. cajennense has at least three hosts during its cycle (Travassos & Vallejo-Freire 1944). A. cajennense have low population densities in birds, possibly due to grooming behavior which may be considered an avian defense against ectoparasites. An evidence of grooming is the fact that ticks are encountered mostly on body parts which birds' beaks cannot reach easily, such as the head and the neck. Non-Passeriformes showed zero prevalence, probably because this group was mostly represented by hummingbirds (Trochilidae) which do not seem to host ticks, as was observed at this study and also by Marini et al. (1996) and Marini and Couto (1997). When exchanging hosts, ticks depend on the frequency of encounter with their hosts (Dobson & May 1986) and the foraging behavior of hummingbirds may reduce the possibilities of such encounters. According to Travassos and Vallejo-Freire (1944) and Serra Freire (1982), A. cajennense is found in its adult phase from late September to late March. Larvae, however, are more frequent during May and June, and nymphs during July and September, with some variations among localities, climate, temperature and hygrometry degree. Here we observed a pattern similar to the one described by the above authors for domestic animals (Figs. 2, 3). Larvae and nymphs of A. cajennense can remain up to 100 days without feeding, maintaining their ability to attack hosts and to complete their life cycle (Olivieri & Serra Freire 1984a, b, Souza & Serra Freire 1992, Serra Freire & Olivieri 1992). This characteristic, associated with the need for three hosts during its evolutive cycle, and their relative readiness to parasitize attacking domestic and wild animals, as well as humans, enable infected individuals to inoculate infectant material during the next phase to another animal or man (Travassos & Vallejo-Freire 1944). When habitats are destroyed, fragmented or modified, different potential hosts make contact with each other, allowing the exchange of their associated arthropods. If these ectoparasites are vectors, the exchange of diseases may occur. Epidemics occur easily when diseases move within new host populations which have not been exposed in their natural habitat (Gettinger & Ernest 1995). The movement of domestic species into wild areas associated with habitat fragmentation increase this risk, which could be kept to a minimum if natural communities were kept as intact as possible, and free of domestic animals. This makes it important to know in greater detail the role of wild species, such as birds, as dispersers of this tick. An integration between ecology and veterinary medicine will allow a better understanding of population dynamics of this tick. ACKNOWLEDGMENTS To the students of the Bird Ecology Laboratory (UFMG) for collaboration in the field work. To Bruce Alexander for English revision. To Paulo R Oliveira for criticisms and useful suggestions. To Cristina MN Lopes and Ramiro Botelho for help in tick identification. To Cemave (Ibama) for banding permission. To "Companhia de Saneamento de Minas Gerais" (Copasa) for permission to work at their properties. This investigation was supported by Fapemig, PRPq and CNPq. REFERENCES Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99060b.jpg] [oc99060c.jpg] [oc99060f.jpg] [oc99060e.jpg] [oc99060d.jpg] [oc99060a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}