 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 94(3) 1999, pp: 367-370 T Cell Response of Asymptomatic Leishmania chagasi Infected Subjects to Recombinant Leishmania Antigens Sérgio Ricardo Costa, Argemiro D'Oliveira Júnior, Olívia Bacellar, Edgar M Carvalho/+ Serviço de Imunologia, Hospital Universitário
Prof. Edgard Santos, Universidade Federal da Bahia, Rua João
das Botas s/no, 40110-160 Salvador, BA, Brasil
Received 28 July 1998
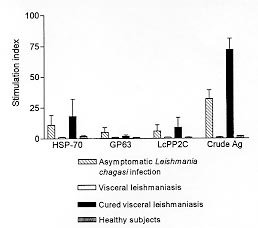
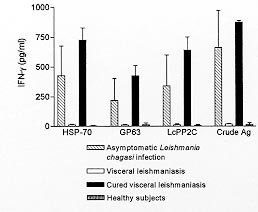
Code Number:OC99068 In areas of Leishmania chagasi transmission the ability to control leishmania infection is associated with IFN-g production. In visceral leishmaniasis down-regulation of T cell responses is mediated by interleukin-10 (IL-10). In this study we evaluated the lymphoproliferative response, IFN-g and IL-10 production on lymphocyte cultures stimulated with recombinant leishmania antigens in subjects with asymptomatic L. chagasi infection. There was a statistically significant difference in the lymphoproliferative response of the subjects with asymptomatic infection as compared to patients with visceral leishmaniasis and healthy subjects with respect to crude antigens (p<0.01), gp-63 (p<0.05) and hsp-70 (p<0.01), as well as between asymptomatic L. chagasi infected subjects and patients with visceral leishmaniasis with respect to the response to all antigens tested. The IFN-g production observed in the group with asymptomatic infection with all the three recombinant antigens tested was higher (p<0.01) than that observed in patients with visceral leishmaniasis and in healthy subjects. Furthermore, in individuals with asymptomatic infection, IL-10 levels in cultures stimulated with recombinant antigens were very low. This study shows that lymphocytes from individuals with asymptomatic L. chagasi infection are able to recognize recombinant leishmania antigens with production of a cytokine that is associated with leishmania killing. Key words: asymptomatic Leishmania chagasi infection - recombinant leishmania antigens - T cell responses Outbreaks of visceral leishmaniasis (VL) have been documented in urban areas of big cities in Central and South American (Grimaldi et al. 1989, Jerônimo et al. 1994). The infection is transmitted by the phlebotomes Lutzomya longipalpis and dogs are considered an important reservoir of Leishmania chagasi. Since control measures such as spread of insecticides and sacrifice of dogs have not been effective in control L. chagasi transmission, immunoprophylaxis has been a goal to control visceral leishmaniasis. Several antigens have been isolated from different leishmania species. These antigens are recognized by serum from VL patients (Schreffler et al. 1993) and by T cells from patients cured of VL or with cutaneous or mucosal leishmaniasis (Reed et al. 1990). However the absence of good experimental model for L. chagasi infection have limited identification of antigens able to induce a protective immune response. IFN-g production is the most important macrophage activating factor to kill leishmania (Murray et al. 1983) and lymphocytes from VL patients which do not produce IFN-g when stimulated by leishmania antigen (Carvalho et al. 1985). In VL down-regulation of T cell response is mediated by interleukin-10 (IL-10) (Ghalib et al. 1993). Therefore in an evaluation of vaccine antigen candidates, these cytokines should be measured. The proposal of the present study was to evaluate whether cells from subjects with subclinical L. chagasi infection recognize recombinant leishmania antigens. Candidate vaccine antigens should hopefully produce immune response similar to those seen in individuals who have asymptomatic infections and are therefore immune. MATERIALS AND METHODS Patients - Monte Gordo is an endemic area of L. chagasi transmission where epidemiological, immunological and clinical studies have been performed in the last five years (D'Oliveira Jr et al. 1997). Serological and clinical surveys are performed annually and seroconverting subjects are considered to be infected with L. chagasi. These individuals lived in the same house or were neighbors of a patient with VL and denied previous history of kala-azar. Follow-up clinical examinations are done every six months to determine signs and symptoms of VL. Participants of this study included ten individuals with asymptomatic L. chagasi infection, six subjects cured of VL, three patients with active VL diagnosed by documentation of amastigote forms of leishmania in Giemsa-stained bone marrow aspirates, and three healthy subjects living outside the endemic area. Subjects with subclinical L. chagasi infection have been followed for more than three years after seroconversion. These individuals remain without clinical evidence of VL but with positive delayed hypersensitive Montenegro test. Subjects cured of VL had the disease three to four years before their participation in the study. Leishmania antigens - As previously described (Carvalho et al. 1985) leishmania lysate (crude antigens) was obtained from promastigotes of a strain characterized by monoclonal antibodies and isoenzymes, as L. chagasi (MHOM Ba 62). The procedures for rgp-63, rhsp-70, LcPP2C production and purification have been previously described (MacFarlane et al. 1990, Russo et al. 1991, Burns et al. 1993). These antigens were generously provided by Dr Reed (Infectious Diseases Research Institute). Lymphocyte blastogenesis assay - Peripheral blood mononuclear cells (PBMC) used for the lymphocyte assays were obtained from heparinized venous blood by centrifugation over lymphocyte separation medium (LSM; Organon Teknika Corporation, Durham, NC). After washing three times in 0.9% NaCl, PBMC were re-suspended in RPMI 1640 culture medium (Gibco, Grand Island, NY) with 200 IU/ml of penicillin and 100 m g/ml of streptomycin. Lymphocyte blastogenesis assays were performed as previously described (Horwitz & Garret 1977). Patients' and controls' PBMC were adjusted to a concentration of 106 cells/ml in RPMI containing 10% normal AB serum. Aliquots of 0.2 ml containing 2x105 cells were cultured in triplicate in microtiter plates (Falcon, Becton Dickinson Co., Oxnard, CA). Cells were stimulated with crude L. chagasi soluble lysate at a concentration of 10 m g/ml. For other antigen cells were stimulated with 100 mg/ml, a concentration previously determined by a dose curve response testing with healthy controls and subjects cured of visceral leishmaniasis cells. Cultures were maintened at 37ºC in 95% air, 5% CO2 for five days. Cells were pulsed with 1 m Ci of [3H] tymidine (6.7 ci/mmol) (New England Nuclear, Boston, MA) during the last 6 hr of culture and collected with Skatron harvester (Flow Laboratories, Rockville, MD). The incorporation of [3H] tymidine was quantitated by liquid scintilation spectometry and results are expressed as mean ± SD of the SI, calculated by dividing the cpm of stimulated culture by cpm of unstimulated culture. Evaluation of IFN-g and IL-10 production - IFN-g and IL-10 levels were determined in supernatants of lymphocyte cultures. Briefly, PBMC (3X106 cells/ml) from patients and controls were stimulated with crude L. chagasi antigen (10 m g/ml), gp-63, hsp-70 and LcPP2C (100 m g/ml) at 37ºC in 95% air, 5% CO2, for 72 hr. After this time, supernatants were collected and the levels of IFN-g and IL-10 were determined by Elisa using a sandwich technique (Russo et al. 1992). A standard curve was used to express the results in pg/ml. RESULTS There was no difference between the groups with respect to age and gender. The lymphopro-liferative response expressed as Stimulation Index to recombinat leishmania antigens (hsp-70, gp63 and LcPP2C) are shown in Fig. 1. There was great variability in the response of the asymptomatic subjects to crude and recombinat leishmania antigens. There was a statistically significant difference in the lymphoproliferative response of subjects with asymptomatic infection as compared to visceral leishmaniasis patients and healthy subjects with respect to crude antigen (p<0.01), gp-63 (p<0.05) and hsp-70 (p<0.01). There was also a statistically significant difference between asymptomatic L. chagasi infected subjects and patients with VL in respect to the response to LcPP2C. While IFN-g production was observed in the group with subclinical infection with all the three antigens tested (Fig. 2), patients with VL and healthy controls did not produce IFN-g in response to crude or recombinant antigens. Levels of IFN-g to hsp-70 (428 ± 253 pg/ml ranging from 162 to 910pg/ml), to LcPP2C (341 ± 263pg/ml ranging from 66 to 971pg/ml) and gp-63 (221 ± 183pg/ml ranging from 8 to 698pg/ml) in supernatants of lymphocyte culture of asymptomatic subjects were higher (P<0.01) than those observed on lymphocyte culture of patients with VL (14 ± 5pg/ml, 16 ± 7pg/ml, 13 ± 3pg/ml respectivelly) and in healthy subjects (< 8pg/ml, 10 ± 17pg/ml and 35 ± 6pg/ml respectivelly). IFN-g production was documented in all ten subjects with asymptomatic L. chagasi infection upon stimulation with Hsp-70 and LcPP2C. The gp-63 induced IFN-g in seven of eight subjects tested. IL-10 production in cultures stimulated with recombinant antigens was very low (hsp-70 17 ± 10pg/ml, gp-63 16 ± 9pg/ml, LcPP2C 13 ± 8pg/ml and crude antigen 28 ± 8pg/ml). There was no difference (P>0.05) in the production of IL-10 in respect to subjects with asymptomatic infection and healthy subjects.
FIGURE 1
FIGURE 2 DISCUSSION In areas of L. chagasi transmission, the majority of the subjects infected by leishmania will develop an asymptomatic or self-healing subclinical L. chagasi infection. The ability to control L. chagasi infection is associated with IFN-g synthesis, and development of disease is characterized by poor IFN-g production (Carvalho et al. 1992). The down regulation of IFN-g production in VL is mediated by IL-10, a cytokine known by its ability to suppress macrophage and lymphocyte function (Carvalho et al. 1994). Although lymphocyte proliferation has been widely used to analyze T cell function, the documentation that the CD4+ population is heterogenous regarding to the cytokine profile secreted, indicates that cytokines should be measured to determine if an immune response can be protective or deleterious. Additionally, lymphocyte proliferation is less sensitive than cytokine production. In the present study, while the 42 kDa protein fraction of L. chagasi (LcPP2C) and the gp-63, an abundant and conserved leishmania surface cell protein, did not induce strong T cell proliferation in subjects with subclinical L. chagasi infection and in individuals cured of VL, the IFN-g production mediated by these antigens was significantly higher than that observed in healthy controls and in patients with VL. Of the recombinant antigens tested the hsp-70 was the one that induced the highest lymphocyte proliferation and IFN-g production. Although hsp-70 is found in other pathogens such as Schistosoma mansoni and Trypanosoma cruzi this antigen did not induce proliferation or IFN-g production in four healthy subjects evaluated. Some of these recombinant antigens have been previously shown to induce lymphocyte proliferation and IFN-g production in subjects cured of visceral and in patients with cutaneous or mucosal leishmaniasis (Russo et al. 1991, Burns et al. 1991). There is no data concerning the ability of these antigens to induce IL-10 and no characterization of the immune response in subjects able to control leishmania infection. Since it has been documented that the same antigen can stimulate cells to produce both IFN-g and IL-10 as well as IL-4 (Skeiky et al. 1995), it is important to determine the cytokine profile produced by PBMC stimulated in vitro by leishmania vaccine candidate antigens. IL-10 has been recognized as the major cytokine that down-regulates IFN-g in L. chagasi infection (Ghalib et al. 1993, Carvalho et al. 1994) and this cytokine was poorly secreted by T cells from resistant subjects infected by L. chagasi when stimulated with recombinant antigens. Although it can be argued that a Th1 type profile was documented in such individuals, as they are prone to a Th1 type of a response secondary to being at the resistant pole to infection, the most important finding was that these individuals recognized recombinant antigens of leishmania with production of cytokine that is associated with leishmania killing. This type of binding leads to the first step of antigen selection in experimental models and in human being, in order to evaluate ability to induce protection against leishmania infection. ACKNOWLEDGMENTS To Dr Steve G Reed for supplying the recombinant antigens, Aurinha Barbosa health assistant at Monte Gordo and Elbe Myrtes S Silva and Jackson L Moreira for computer assistance. This work was supported by NIH grant AI-30639, UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR) 940687, Programa de Núcleo de Excelência and Financiadora de Estudos e Projetos. REFERENCES Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99068a.jpg] [oc99068b.jpg] |
| |||||||||

{kind=link}
{kind=link}