 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 94(3) 1999, pp: 387-394 Increased Pro-inflammatory Cytokines (TNF- a and IL-6) and Anti-inflammatory Compounds (sTNFRp55 and sTNFRp75) in Brazilian Patients during Exanthematic Dengue FeverLuzia MO Pinto, Solange A Oliveira*, Elzinandes LA Braga, Rita MR Nogueira, Claire F Kubelka/+ Departamento de Virologia, Instituto Oswaldo Cruz, Av. Brasil
4365, 21045-900 Rio de Janeiro, RJ, Brasil Received 1 September 1998
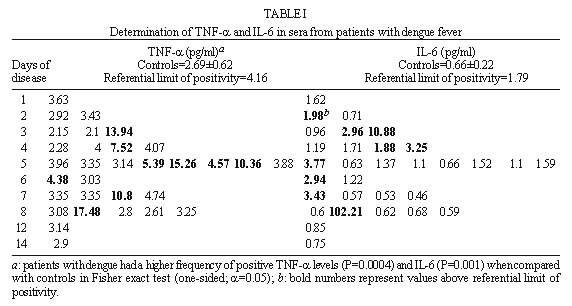
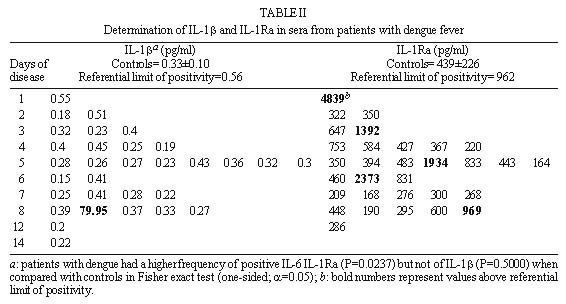
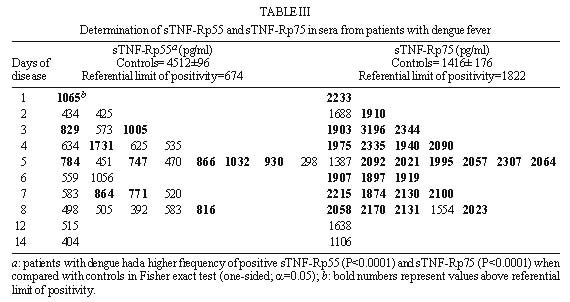
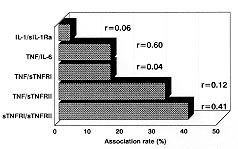
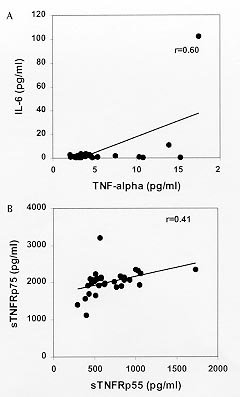
Code Number:OC99072 Pro-inflammatory cytokines, tumor necrosis factor (TNF-a ), interleukin-6 (IL-6) and interleukin-1b (IL-1b ) as well as anti-inflammatory compounds, soluble TNF-Receptor p55 (sTNFRp55), sTNFRp75 and IL-1 receptor antagonist (sIL-1Ra), were investigated in 34 Brazilian cases of dengue fever (DF) originated from a study of exanthematic virosis. The presence of pro-inflammatory cytokines was detected in sera from these patients by ELISA. TNF-a and IL-6 levels were significantly higher than control subjects in 32% and 52% patients, respectively. To our knowledge this was the first time a receptor antagonist and soluble receptors for cytokines were detected in sera obtained during exanthematic DF without hemorrhagic manifestations. Both sTNFRp55 and sTNFRp75 were consistently elevated in 42% and 84% patients, respectively. Most patients had IL-1b levels not different from those of normal subjects, except for one case. Only 16% patients had altered levels of IL-1Ra. Previous studies in dengue hemorrhagic fever patients demonstrated production of these soluble factors; here we observed that they are found in absence of hemorrhagic manifestations. The possible role of these anti-inflammatory compounds in immune cell activation and in regulating cytokine-mediated pathogenesis during dengue infection is discussed. Key words: dengue - tumor necrotis factor a - interleukin 6 - soluble tumor necrosis - factor receptor Dengue disease is caused by single-stranded positive sense RNA arboviruses, belonging to the Flaviviridae family and classified as four antigenically distinct dengue virus serotypes (Sabin & Schlesinger 1945, Hamon et al. 1960, Brown 1986). Infection can be asymptomatic or lead to different forms of disease. Dengue fever (DF) is characterized by mild systemic manifestations such as fever, retro-orbital headache, severe myalgias, and rash. Some patients develop a more severe and life-threatening syndrome termed dengue hemorrhagic fever (DHF) where plasma leakage takes place into interstitial spaces, resulting in hypovolaemia, thrombocytopenia and hemorrhage; circulatory collapse leading to shock may occur and is then referred as dengue shock syndrome (DSS) (Halstead et al. 1988, Halstead 1990, PAHO 1994). The worldwide number of annual DF cases is estimated to be more than 100 million, with 250,000 reported cases of DHF (Monath 1994). In Brazil, about 400,000 cases of dengue were notified this year during the first six months (Brazilian National Health Foundation, Heath Ministry 1998). The pathogenesis of dengue disease is not fully understood and is considered an immunopathologic process associated with prior immune sensitization by a heterotypic virus (Halstead 1980, Monath 1986, Kurane et al. 1991). Dengue infection provides lifelong homotypic immunity, but only transient cross-protection against other serotypes is achieved, making sequential infection possible (Pang 1987, Kliks et al. 1989). The relative risk of experiencing most severe forms of disease has been considered to be several fold higher after secondary infection (Monath 1986). Mononuclear phagocytes appear to be principal target cells for dengue virus replication (Halstead 1980, Anderson et al. 1997). The underlying mechanism is believed to involve activation of virus infected macrophages and producion of cytokines such as tumor necrosis factor-a (TNF-a ), interleukin-6 (IL-6) and interleukin-1b (IL-1b ) (Yang et al. 1995, Hober et al. 1996b). These pro-inflammatory cytokines are associated in vivo with an acute phase response, and may result in liberation of chemotactic peptides, fever and activation of endothelial cells leading to vascular permeability which may be the phenomenon involved in the pathogenesis of dengue infection (Anderson et al. 1997). The production of pro-inflammatory cytokines (TNF-a , IL-6 and IL-1b ) and the extent of the inflammatory response are partially modulated by anti-inflammatory compounds. Soluble extracellular domains of 55-kDa and 75-kDa, sTNFRp55 and sTNFRp75, are liberated extracellularly from membrane-bound TNF receptors (Kornelisse et al. 1996). IL-1 receptor antagonist (IL-1Ra) may function as an inhibitor by antagonizing IL-1 binding to IL-1 cell surface receptor in a competitive interaction (Pereda et al. 1995). Several cytokines were detected in sera from patients undergoing viral infections. Their exacerbated induction is associated with pathological changes such as hepatic lesions in acute (Torre et al. 1994) and chronic (Yoshoka et al. 1989) viral hepatitis or HIV-1 replication in AIDS (Poli & Fauci 1993). They play also a role in septic shock which has some clinical features similar to DSS (Dinarello 1996a). Pro-inflammatory cytokines were found to be increased in Asiatic and American patients with DHF/DSS (Hober et al. 1993, 1998, Kuno & Bailey 1994, Iyngkaran et al. 1995, Kubelka et al. 1995) and recently soluble TNF receptor was detected in DHF/DSS patients (Hober et al. 1996b, Bethell et al. 1998). Nevertheless, these investigations are far from elucidating the complex mechanisms of immunopathology during dengue disease; more cases in different countries and several immunological parameters deserve to be studied. Moreover, none of these works discuss the cytokine profile for DF. During the present work we observed the presence of pro-inflammatory cytokines (TNF-a , IL-6 and IL-1b ) and anti-inflammatory compounds (sTNFRp55, sTNFRp75 and sIL-1Ra) in sera of Brazilian patients with exanthematic DF without hemorrhagic manifestations and infected with Dengue-1 or -2 viruses. MATERIALS AND METHODS Patients and laboratory diagnostics - Thirty four cases of DF included in this work originated from a study on exanthematic virosis with patients attended at the Antônio Pedro University Hospital, Niterói in 1995-1996 (22 women, 12 men; 1-58 years old, average 29 ± 14.9). All patients were diagnosed based on clinical manifestations of DF and confirmed serologically by the presence of IgM in MAC-ELISA test (Nogueira et al. 1992). Also, all patients were seronegative for measles and rubeolla specific IgMs. Hemagglutination inhibition test (HI) (Clarke & Casals 1958) was carried out and titers equal or greater than 1/160 were considered as secondary infection for a post-epidemic period. Nine healthy individuals (3 women, 6 men; 22-45 years old, average 32.4 ± 7.9) were used as negative controls. Cytokine and receptor cytokine assays - Serum samples were obtained during appointment and were stored in aliquots at -20°C until use for different assays. Serum levels of TNF-a , IL-6 and IL-1b were assayed in High Sensitivity ELISA kits (QuantikineTM HS, R&D Systems) and sTNFRp55, sTNFRp75 and IL-1Ra in ELISA kits (QuantikineTM, R&D Systems) according to the manufacturer's instructions. They presented the following limits of sensitivity achieved in standard curves: TNF-a , 0.5 pg/ml; IL-6, 0.156 pg/ml; IL-1b , 0.125 pg/ml; sTNFRp55, 7.8 pg/ml; sTNFRp75, 7.8 pg/ml and IL-1Ra, 46.9 pg/ml. Statistical analysis - Statistical analysis was performed by calculating a t=2.306 value, found in the table of percentage points of the t Distribution: n =8 is the number of control samples (9) minus 1 and a =0.025 is the degree of significance used for the test. A referential limit value for positivity was calculated according to the following formula: Average of values from control samples + [Standard Deviation of values from control samples X t(n=8;a=0.025)]. Determinations above referential limit values were considered positive. The correlation coefficient (r) was calculated between levels of different factors. The Fisher exact test was applied to determine if the frequency of positive patients for circulating soluble factors was significative and to associate arthropathy or the type of infection (primary/secondary) with the production of soluble factors. A p value of £ 0.05 was required for differences to be considered significant. RESULTS Clinical manifestations and laboratory features - All 31 patients showed always fever, rash and no hemorrhagic manifestations, except for one case without fever and three others who exhibited discrete petechia. Seventeen patients, all adults, had arthritis and/or arthralgia, besides usual dengue clinical manifestations (malaise headache and retroorbital pain, myalgia, anorexia, nausea, vomiting). Fifteen out of 28 patients tested for HI had primary infection and 13 had secondary. Viruses isolated in the State of Rio de Janeiro during the year 1990 and onwards were serotypes 1 and 2. During the years 1995-1996 Dengue-1 and Dengue-2 co-circulated in Niterói, but serotypes of individual sera were not tested. Pro-inflammatory cytokines levels - The TNF-a concentration in plasma was increased in one third (10 out of 31) of tested patients with DF (Table I), in comparison with healthy individuals. The maximal value achieved was 17.5 pg/ml, in an adult patient on day 8 of disease. Approximately half the patients (16 out of 31) showed a rise in levels of IL-6 ( Table I). The maximal value was 102 pg/ml in the same patient whose value for TNF-a was the highest. Only one adult patient out of 31 was positive for IL-1b and exhibited a high level of the cytokine, 80 pg/ml (Table II). Again this was the same patient who had high levels of TNF-a and IL-6. Soluble receptor and receptor antagonist levels - Five patients out of 31 (16%) showed increased levels of sIL-1Ra ( Table II). The maximal value detected was approximately 5 ng/ml, in a patient during the first day of disease. Among all factors sTNFRp75 was found at highest frequency, 84% (26 of 31), in the sera of dengue patients. The concentration of sTNFRp55 in plasma was elevated in 42 % (13 of 31) DF ( Table III). Age-dependent incidence of soluble factors - Pro-inflammatory cytokines and anti-inflammatory compounds were present in children as well as in adults. Due to the low number of child patients (seven), though, no statistics was performed among age groups. Association and correlation between soluble factors present in serum from dengue patients - Soluble receptors for TNF-a (sTNFRp55 and sTNFRp75) appeared simultaneously in 12 out of 30 patients studied. The association of TNF-a with sTNFRp75 wase more frequent than TNF-a and sTNFRp55. Concomitant TNF-a and IL-6 was observed in only six patients (Fig. 1).
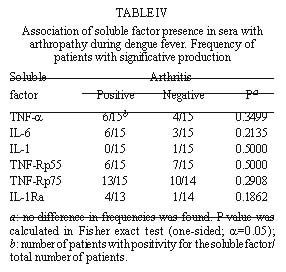
FIGURE 1 If the correlation coefficient is calculated, TNF-a and IL-6 show a significant correlation; a weaker correlation was also found for sTNFRp55 and sTNFRp75 (Fig. 2). Other factors could not be associated.
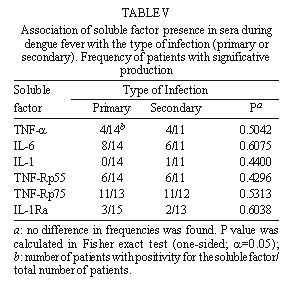
FIGURE 2 In the only exception for a positive IL-1b , the patient had levels 255-fold higher than the average of the rest of the patients and no sIL-1Ra was produced in this serum. As mentioned before this patient had the highest values for the three cytokines, but clinical manifestations remained without outstanding features. All five patients positive for sIL-1Ra produced no IL-1b ; thus, no association between these two factors was shown. Association of soluble factor production with arthropathy or to the type of infection (primary or secondary) during dengue fever - Attempts to associate arthritis and/or arthralgia with pro-inflammatory cytokines or with anti-inflammatory compound production were not successful ( Table IV). No significance was found using Fisher exact test in data from all factors measured. Furthermore, no statistical difference was observed between groups of primary and secondary infection in the production of these soluble factors ( Table V). DISCUSSION Dysregulated expression of inflammatory cytokines TNF-a , IL-6 and IL-1b is known to be implicated in immunopathologic mechanisms such as inflammation, necroinflammatory injuries and endotoxic shock (Dinarello 1996a). Viral genic products originating from HTLV-I, Hepatitis B Virus are able to activate promoters from transcription factors such as NFk B and NF-IL-6 (Kishimoto et al. 1994) known to induce pro-inflammatory cytokines which are frequently detected circulating during viral infections (Hober et al. 1989). During this work, TNF-a , IL-6, sTNFRp55 and sTNFRp75 were detected in the sera of Brazilian patients with DF, devoid of hemorrhagic manifestations and infected with Dengue-1 or -2. In general, the levels of cytokines in most of our DF patients were lower than those described in literature for DHF/DSS (Hober et al. 1993, 1996a, Kuno & Bailey 1994, Iyngkaran et al. 1995, Kubelka et al. 1995, Bethell et al 1998). This may be explained either by relative disease mildness, virus load, by differences in viral serotype, host genetic factors that may influence in the immunological responses or in test sensitivity. Our results reveal that cytokines, TNF-a and IL-6 and soluble TNF receptors were present in significantly increased levels until the eighth day of DF, in several but not in all patients. Factor production at later stages of disease remains to be determined. The levels of TNF-a observed suggest a production from the third day of disease onwards. IL-6 was already present at significant levels at patients at the first two days and sTNFRp75 appeared in the first day and could be detected in most patients (four out of five) at the eighth day. Nevertheless, higher number of patients should be tested to confirm these results. In contrast, IL-1b seems not to be altered as DF progresses. This is in agreement with earlier studies on DHF/DSS (Hober et al. 1993). Circulating IL-1b levels detected usually in pathologic conditions are relatively low compared with levels of IL-6 and TNF-a . Unlike TNF-a , IL-6 or IL-1Ra, a significant amount of proIL-1b remains inside the cell. IL-1b also binds to large proteins such as a2-macroglobulin, complement, and the sIL-1RII, which in turn binds preferentially to IL-1b when compared to IL-1a or IL-1Ra (Dinarello 1996a), being not easily reactive in regular ELISA assays. The biological properties of TNF-a share remarkable similarities to those of IL-6 and IL-1b : they are endogenous pyrogens and inducers of acute-phase responses. IL-1-b and TNF-a induce IL-6 production. It has been claimed that levels of IL-6 often may better correlate with severity of an infectious disease (Dinarello 1992, 1996b). In our work IL-6 was the most frequently detected cytokine. Moreover, one patient with all three cytokines elevated had levels of IL-6 six-fold higher than TNF-a and 1,5-fold higher than IL-1b . Previous work studying DHF patients described contradictory data relating IL-6 levels and severity of disease (Hober et al. 1993, Kuno & Bailey 1994, Bethell et al. 1998). Exanthema is not exclusively dependent on the elevation of circulating factors, since all patients developed rash and some were negative for their production; on the other hand, hemorrhagic manifestations cannot be directly associated with production of any of the circulating factors. Moreover, the presence of cytokines in circulation seems not to be related to arthropathy. During malignant ascites (Van Zee et al. 1992) the concentration of sTNFR in plasma was lower than in the synovial fluid where inflammation actually occurred; moreover, differences in factor localization were correlated with disease severity. If cytokines and/or soluble receptors play some role in joint manifestations during dengue disease, local production should be investigated. An association found between two soluble TNF receptors (sTNFRp55 and sTNFRp75) could indicate that their induction mechanisms might be related. Circulating sTNFRs may provide different regulatory pathways for modulating TNF-a effects. sTNFRs can compete for TNF-a with cell surface receptors and thus reduce the activity of the cytokine; on the other hand they may also enhance TNF-a function regulating its bio-availability, most likely by stabilizing the active TNF-a oligomer (Leuwenberg et al. 1994). According to our data TNF-a seems to have a better association with sTNFRp75 than p55. If this is confirmed, it may be postulated that sTNFRp75 might modulate TNF-a activity in vivo during DF. Furthermore, we observed in both normal and DF sera that sTNFRp75 was more abundant than sTNFRp55; this is in accord with usual descriptions from in vivo studies during inflammatory processes or in normal individuals (Hart et al. 1996). IL-1Ra is stimulated under conditions where pro-inflammatory cytokines would be inhibited. IL-10 and IL-4 suppress macrophage release of pro-inflammatory cytokines (TNF-a , IL-6 or IL-1b and IL-8) and stimulate the secretion of IL-1Ra and nonsignaling type II IL-1 receptor (Tilg et al. 1997). Also, IL-4 inhibits the release of sTNFRs from monocytes. Therefore, besides soluble receptors, other cytokines can antagonize the biological activity of pro-inflammatory cytokines and could eventually be acting during dengue infection. This could explain IL-1Ra has increased expression in five patients but attempts to inversely correlate with pro-inflammatory cytokines have failed until now. Moreover, in an earlier work, Hober et al. (1996a) described that in Dengue-3-induced DHF/DSS no apparent correlation between TNF-a and sTNFRp75 could be made. Taken together, clinical observations and serum titration described here show that pro-inflammatory cytokines (such as TNF-a and IL-6) are also produced during mild non hemorrhagic manifestations of exanthematic DF and it is likely that they play a role in this pathology, as described earlier for DHF/DSS. Severity may be related to the amount of circulating cytokines. Soluble receptors, mainly sTNFRp75, are also increased and, considering its association with TNF-a , may be used as a marker of immunological activation, since it is more stable than pro-inflammatory cytokines. From our findings the question still remains: could sTNFp75 act as modulator of an excessive biological activity of TNF-a (Tilg et al. 1997) preventing severe disease during DF? Further studies deserve to be performed to broaden our understanding about the balance among different circulating immunological factors and their effects in development of disease. Acknowledgments To Drs Takumi Iguchi and José A Losana for statistical advice, Drs Hermann G Schatzmayr and Marilda M Siqueira for continuous encouragement, Dr Marize P Miagostovich and Ms Eliane Saraivo for performing laboratory diagnostics. Financial support: Fiocruz, CNPq and Colab, Brazil. REFERENCES
Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99072e.jpg] [oc99072b.jpg] [oc99072d.jpg] [oc99072a.jpg] [oc99072g.jpg] [oc99072c.jpg] [oc99072f.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}