 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 94(4),Jul./Aug. 1999: pp Density of Sand Flies (Diptera: Psychodidae) in Domestic and Wild Animal Shelters in an Area of Visceral Leishmaniasis in the State of Rio Grande do Norte, Brazil Maria de Fátima Freire de Melo Ximenes/+, Maria de Fátima de Souza, Eloy Guilhermo Castellón* Departamento de Microbiologia e Parasitologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, 59072-970 Natal, RN, Brasil *Coordenação de Pesquisa em Entomologia, Instituto Nacional de Pesquisa da Amazônia, Manaus, AM, Brasil + Corresponding author. Fax: +84-211.9210. E-mail: ximenes@cb.ufrn.brReceived 2 March 1998
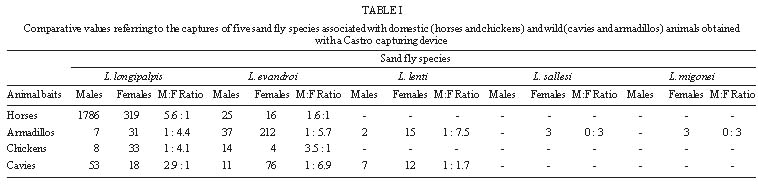
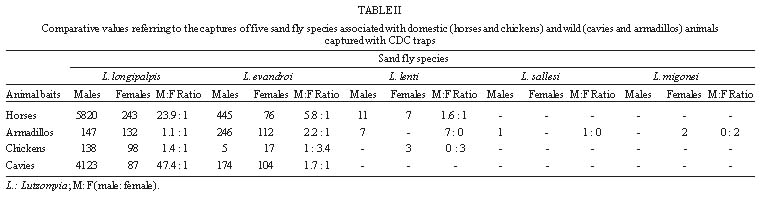
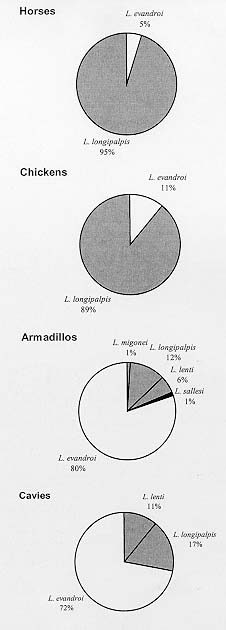
Code Number:OC99080 The objective of the present study was to determine the association of sand flies with the presence of domestic and wild animals in the peridomiciliary area. The sand flies were collected using direct aspiration and CDC light traps placed in animal shelters. The results suggest that different sand flies species have different behavioral characteristics in an apparent preference for animal baits and that Lutzomyia longipalpis and Lu. evandroi were the most eclectic species regarding their biotope choice. Lu. longipalpis showed a distinct preference for horses and Lu. evandroi for armadillos. Key words: Lutzomyia longipalpis - Lutzomyia evandroi - host preference - Rio Grande do Norte - Leishmania A wide variety of animal aggregation is observed in several animal orders, involving some environmental adaptation that will ensure permanent or temporary cohesion among groups of the same species. Thus, these aggregations correspond to groupings of individuals around the same stimulus or resource (Carty 1980, Deag 1981). Studies on the aggregation behavior of dipterans (Nematocera) are scarce, although their importance is recognized for the understanding of questions that cannot be resolved solely on the basis of physiological mechanisms. Little is known about the behavior of sand flies, including the vectors of the different species of Leishmania. An understanding of the behavior of these insects in nature is important in the design of control measures. The selection of an appropriate site for oviposition, the search for food sources and resting sites by females are behavioral responses that affect the reproduction and population density of the species. Female sand flies require vertebrate blood obtained by single or multiple meals for the maturation of their eggs. Thus, depending on their degree of adaptation to sites disturbed by humans, some species are more easily detected in the peridomiciliary environment than others. On this basis, the food sources and shelter and resting sites for these species vary according to the presence of domestic and wild animals in the peridomiciliary area. Studies on Lutzomyia longipalpis (Dye et al. 1991, Ward et al. 1993, Quinnell & Dye 1994, Kelly & Dye 1997) have shown the importance of semiochemi-cals (pheromones and host - produced kairomones) for the aggregation of sand flies in the peridomi-ciliary environment. Human leishmaniasis represent an important public health problem in the Northeast region of Brazil. In Rio Grande do Norte, the number of cases of visceral leishmaniasis reported to the National Health Foundation is higher than that of tegumentary leishmaniasis. It is known that the population density of Lu. longipalpis, which transmits Le. chagasi, is affected by peridomiciliary conditions and it is often associated with the presence of domestic animals (Forattini 1960, Sherlock & Guitton 1969). In this region, including the rural areas of Rio Grande do Norte, dwellings are often located closer to native vegetation where small mammals such as marmosets (Callithrix jacchus), opossums (Didelphis) and rodents are frequently found. In addition, cavies and armadillos may be captured and reared for some time for later use as food. The objective of the present study was to investigate the behavioral aspects involved in the relationship between Lutzomyia species and different animals and its possible effect on the occurrence of visceral leishmaniasis in the study area, in view of the importance of the association of vectors with animals present in the peridomiciliary area. Study area - Nísia Floresta, one of the municipalities with reporting cases of visceral leishmaniasis in Rio Grande do Norte, is located in the oriental littoral zone, 43 km distant from Natal, capital of the state. The climate of this area (elevation 20 m.a.s.l.) is subhumid, with a mean annual rainfall of 1,400 mm, a mean temperature of 26oC and a mean relative humidity of 76%. The estimated population of Nísia Floresta is 15,624 inhabitants and 15 cases of visceral leishmaniasis were reported to the National Health Foundation between 1991 and 1996. The site of studies is located in a farm where the family house is in close proximity of trees as cashew nut tree (Anacardium occidentale), mango trees (Mangifera indica), coco nut (Coccos nucifera) and "mangaba" tree (Hancornia speciosa). Eighty meters from the house there was a small patch disturbed secondary Atlantic forest where marmosets, opossums and rodents were found. MATERIALS AND METHODS Twelve sandfly collections were performed in a site bordering a small forest area near the Bonfim Lagoon using Castro capturing devices and light traps (CDC) in the peridomiciliary environment between October 1996 and May 1997. Manual captures were performed by four people at the same locations and at the same time between 18 and 20 hr in man-made shelters for domestic animals (horses and chickens) and wild animals (cavies Galea spixii) and (armadillos Euphractus sexcintus). Three of these four persons participated in capturing sand flies on animals (horses, chickens, armadillos and caves) and on the walls of armadillo and cavy shelters located in the backyard of the dwelling. In the late afternoon, the horses were brought to a wire enclosure measuring approximately 5 x 5 m. During the day, this enclosure is protected by the shade of a native tree (Protium braziliense) and of a palm (Coccos nucifera). Chickens ranged free during the day and sheltered at night in a chicken coop made of sticks and palm straw, measuring approximately 1.7 x 1.2 m. The armadillos were kept in a brickwork tank measuring approximately 1.2 x 1.2 m, covered with plastic and planks, and the cavies were reared in a brick enclosure partially covered with shingles and planks measuring 1 x 0.7 m. Light traps were set in the shelters of these animals on days when no manual captures were performed, between 18 and 6 hr. RESULTS Captures conducter either using manual capturing devices or CDC light traps yielded 14,690 specimens of Phlebotominae, of which 13,067 were males (Lu. longipalpis, 92.5%; Lu. evandroi, 7.3%; Lu. lenti, 0.2%; Lu. sallesi, 0.01%) and 1,706 females (Lu. longipalpis, 56.3%; Lu. evandroi, 36.2%; Lu. lenti, 2.2%; Lu. migonei, 0.3%; Lu. sallesi 0.2% and Lu. sp., 4.8%). Figure shows the frequency of distribution of manually captured females associated with distinct animals. Ninety five per cent of all sand flies collected on horses were Lu. longipalpis and 80% of all sand flies collected on armadillos were Lu. evandroi. The sex ratio for Lu. longipalpis on horses showed a larger number of males than females, both in manual and light traps captures. For armadillos and chickens, an inverse relationship was observed in manual captures, i.e. one male to four females, while in cavies the number of males was higher than the number of females for the two forms of capture (Tables I, II). For the species Lu. evandroi, the number of males was higher than females in manual captures on horses and chickens ( Table I), whereas on armadillos and cavies this relationship was inverted, with approximately six females to one male. This species, when captured by light trap, showed a larger number of males on horses, armadillos and cavies ( Table II). The number of other Lutzomyia specimens was very small compared to Lu. longipalpis and Lu. evandroi. DISCUSSION The presence of large numbers of sand fly vectors, in peridomiciliary environments (Forattini 1953, 1960, 1976, Deane & Deane 1962, Sherlock & Guitton 1969, Gomes et al. 1983, Brazil et al. 1991, Brandão-Filho et al. 1994, Azevedo et al. 1996, Sherlock 1996) increases the risk of Leishmania transmission at these sites. The seasonal fluctuation of Lu. longipalpis in Bahia was associated with the increased frequency of Didelphis albiventris in the peridomiciliary area and with new cases of visceral leishmaniasis (Sherlock 1996). Domestic and wild animals present in the peridomiciliary area probably attract a large number of sand fly species, thus contributing to the increased density of leishmaniasis vector species in some areas of the state. Rearing of these animals, together with poor hygienic conditions, creates a habitat favoring the aggregation of Lu. longipalpis and of other sand flies. With respect to tegumentary leishmaniasis, we captured Lu. migonei, a species possibly involved in the transmission of Le. braziliensis in Ceará (Azevedo & Rangel 1991) and other regions of Brazil. In southern Brazil it has been shown that domestic animals strongly attract sand flies and that Lu. migonei and Lu. whitmani have different food preferences (Teodoro et al. 1993). In the current study we observed that the species present in the peridomiciliary area probably use the shelters of wild animals as breeding sites and resting places. We observed engorged females resting on the walls of the shelters during the observation period. Although we did not perform a systematic study of the physiological age of females and did not look for eggs or larvae at the site, we observed blood and/or eggs inside most females. These findings led us to assume that the females present at these sites were nullipars, resting after a blood meal, and gravid females, searching for protection from possible predation and environmental pressure as rain and wind. The absence of wind, the humidity, the type of soil and the decomposing organic matter found in armadillo shelters seem to contribute to the presence of the five species captured at these sites. The odor of organic matter attracts gravid females and in addition food extracts and rabbit feces combined with oviposition pheromones elicit a positive response from gravid Lu. longipalpis females (McCall & Cameron 1995). In general, the number of males present in the cavy and armadillo shelters was small compared to the number of females. Although no female pheromones are known, we can not rule out the possibility that females use of some sort of visual or auditive communication to attract males to cavies and armadillos shelters (Ward et al. 1988). The species Lu. longipalpis and Lu. evandroi proved to be eclectic in their choice of biotopes. Lu. longipalpis presented the following decreasing order of preference with respect to animal shelters: horse, armadillo, chicken and cavy; Lu. evandroi: armadillo, cavy, horse and chicken. Horses attracted Lu. longipalpis much more than other animals, perhaps because of the result of their large size and of the degree of adaptation of this sand fly species to the peridomiciliary area, where horses and donkeys are frequently found. The attraction of Lu. longipalpis for humans has been related to body size (Quinnell et al. 1992). However, in another study carried out in the laboratory, it was suggested that this species is opportunistic and is not highly attracted to humans or dogs (Morrison et al. 1993). Similar studies conducted in a field site, also showed a clear preference for chickens by sand flies (Aguiar et al. 1987). Although horses, cavies and armadillos have not been incriminated as reservoirs of Le. chagasi, the presence of these animals in the peridomiciliary area in association with dogs may increase the risk of Le. chagasi transmission since they attract sandflies that may take blood meals from dogs and humans, thus transmitting the parasite. Lu. evandroi was the second most frequently encountered species. It is anthropophilic but has not yet been demonstrated to be a vector for human leishmaniasis. The sex ratio found for Lu. longipalpis in the present study was similar to other reports (Dye et al. 1991, Quinnell & Dye 1994). They consider this finding as a strategy used by the species which is caused by the production of male pheromones and as a consequence females are attracted and other males are recruited to feeding and mating sites (Dye et al. 1991, Quinnell & Dye 1994). At first the males seem to be attracted by the odor of host animals and then the pheromone produced by the first male may act as an additional attractant for male and female sand flies (Quinnell & Dye 1994). The manual captures on horses collected Lu. longipalpis (95.2%) and Lu. evandroi (4.8%) and we frequently observed sand fly copulation. It is possible that, with the presence of a large number of males, those who succeed in mating with feeding females or close to the blood meal source benefit, with a consequent increase in the reproductive success of the species. Lu. longipalpis males were more frequently captured than females using both forms of captures (Tables I, II), with the exception of manual capture on chicken and armadillos. However, the number of sand flies captured in chicken was small when compared to other animals. It is possible that the armadillo shelters are being used as a resting place for engorged and/or gravid sand flies. For Lu. evandroi, the number of males was higher than the number of females in manual captures on horses and chickens, whereas on armadillos and cavies this relationship was inverted (Table I). Lu. evandroi also showed a disproportionate ratio of males to females when captures were made on horses, armadillos and cavies with CDC traps. However, the studies conducted on this species do not permit, as yet, to draw conclusions similar to those obtained for Lu. longipalpis, although Lu. evandroi males have pheromone disseminating structures in tergites (Ward et al. 1993). A higher number of females of Lu. longipalpis or Lu. evandroi captured manually on chicken, cavies and armadillo might indicate that female sand flies use different animals for their meals and they rest in more enclosed protected site as armadillo and cavies shelters in the evening after feeding and/or copulation. In some occasions, we examined the animals shelters during the day and we found few females resting in armadillo shelters. It is possible that during the day, there are other more protected shelters sand flies in the native vegetation found in close proximity of the house for both male and females sand flies than the shelters examined, once these sites are frequently disturbed during the day by people who live and work in the farm. The dinamic involved in sand fly aggregation and animal host is not completely understood. Our findings raises two possible explanations for the aggregation. Animals seem to attract sand flies to the peridomicialy environment as shown by the decrease in sand flies density after removal of domestic animals, rodents and armadillos without any insecticide spraying. Furthermore, it is possible that male Lu. longipalpis utilize hot blood animals (horses) as lekking sites as a result of the species adaptation to more opened places, where Lu. longipalpis can be frequently found. The animal body heat can be important for the spread of pheromone and attraction of sand flies. Horses with the body size and decrease of activity in the evening could be contributing to the perfection of Lu. longipalpis strategy for mating. Finally, males and females aggregate close to the host where they copulate followed by female feeding on the host and/or posture in the more humid and rich organic debris. Despite the availability of behavioral studies on Lu. longipalpis and the attempts to understand the participation of semiochemicals as mediators of the presence of vectors in the human environment, the dynamics of sand fly aggregation in shelters for domestic and wild animals needs further elucidation. In addition, the results obtained in several studies on zoophilic and anthropophilic behavior in a natural environment are conflicting. Thus, an understanding of the relations between sand flies and domestic and wild animals reared by man coupled with the understanding of genetic and behavior differences among the vector population from distinct geographic areas may contribute to improve measures for the control of leishmaniasis in rural areas of the Northeast region, where drought and scarcity of food affect customs and expose man to risks of developing diseases such as visceral and cutaneous leishmaniasis. Acknowledgments To Mr Manoel Silva and his family for allowing the conduction of this study in their property; Dr Selma MB Jerônimo for suggestions; Dr Adalberto Varela for helpful information on the fauna of the State of Rio Grande do Norte; Mr Edson Santana, Ms Janaína Cunha Maciel and Mr Gilvan Gurgel de Albuquerque for help in conducting the sand flies captures. REFERENCES Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99080b.jpg] [oc99080c.jpg] [oc99080a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}