 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 94(4),Jul./Aug. 1999: pp 527-530 Research Note Heterologous Expression of a Trypanosoma cruzi Surface Glycoprotein (gp82) Indicates that Requirements for Glycosylphosphatidyl-inositol Anchoring are Different in Mammalian Cells and this Trypanosome Marcel I Ramirez, Silvia B Boscardin, Rita C Ruiz, Sang W Han*, Glaucia S Paranhos-Baccala**, Nobuko Yoshida, Renato A Mortara, José Franco da Silveira/+ Departamento de Microbiologia, Imunologia e Parasitologia
*Departamento de Biofísica, Escola Paulista de Medicina,
Universidade Federal de São Paulo, Rua Botucatu 862,
04023-062 São Paulo, SP, Brasil **Unité Mixte 103
CNRS-bioMérieux, École Normale Supérieure de
Lyon, 46 allée d'Italie, 69364, Lyon Cedex 07, France Received 30 November 1998 This work was supported by grants from Fapesp, Finep/BID (66/96/0792/00) and PADCT/CNPq (Brazil), International Atomic Energy Agency (8334/RB) and Cyted (Ibero American Project of Biotechnology, Subprogram III, Spain).
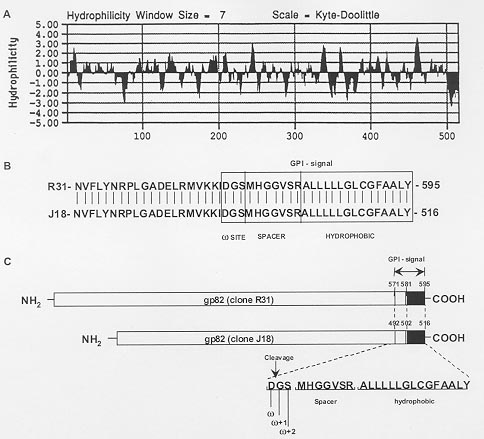
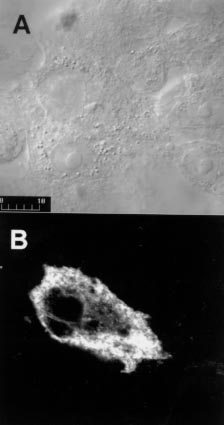
Code Number:OC99097 Key words: Trypanosoma cruzi - surface glycoprotein - heterologous expression - mammalian cells - membrane protein processing - glycosylphosphatidylinositol attachment RESEARCH NOTE Metacyclic trypomastigotes of Trypanosoma cruzi express a stage-specific surface glycoprotein of 82 kDa (gp82) which has been implicated in the invasion of the mammalian host cells (MI Ramirez et al. 1993 Infect Immun 6: 3636-3641, FR Santori et al. 1996 Mol Biochem Parasitol 78: 209-216). Gp82 can induce Ca2+ signal in target cells (ML Dorta et al. 1995 Mol Biochem Parasitol 73: 285-89, RC Ruiz et al. 1998 Biochem J 330: 505-511), an event essential for T. cruzi internalization (I Tardieux et al. 1994 J Exp Med 179: 1535-1540, SNJ Moreno et al. 1994 J Exp Med 180: 1535-1540, Dorta et al. loc. cit.). Structurally, gp82 is a surface glycoprotein containing N-linked oligosaccharide side chains and anchored to the membrane through a glycosylphosphatidylinositol (GPI) moiety (Ramirez et al. 1993 loc. cit., ML Cardoso de Almeida & N Heise 1993 Biol Res 26: 285-312). A cDNA recombinant clone (clone J18, GenBank L14824) encoding gp82 was isolated and characterized (JE Araya et al. 1994 Mol Biochem Parasitol 65: 161-169). It has an open reading frame that encodes a polypeptide of 516 amino acids with a molecular mass of 55.6 kDa, which may correspond to the precursor of gp82 devoid of N-linked oligosaccharides (Araya et al. loc. cit.). Hydrophobicity analysis of gp82 indicates the presence of a nonpolar region of 15 amino acids at the extreme carboxy-terminus which is characteristic of proteins anchored to the cell surface by a GPI linkage (Fig. 1). Although many features of the biosynthesis of surface membrane proteins have been conserved throughout evolution, the requirements for translocation of the nascent peptide chain across the endoplasmic reticulum membrane and GPI anchor attachment may be not the same in mammals and parasitic protozoa. Previous studies have indicated that the GPI signal of the variant surface glycoprotein of T. brucei is non-functional in the mammalian systems (P Moran & IW Caras1994 J Cell Biol 125: 333-343). They showed that the cleavage/attachment site of the variant surface glycoprotein GPI signal was incompatible with the mammalian machinery for GPI attachment. It is of great interest to extend this analysis to T. cruzi, and to study whether the requirements for GPI attachment are the same in parasitic trypanosomes. The goal of the present work was to test whether the gp82 signals for GPI anchor cleavage/attachment function in mammalian cells. The gp82 of T. cruzi was expressed in Vero cells transfected with pcDNA3 plasmid carrying the gene gp82. The complete open reading frame of gp82 gene was subcloned into pcDNA3 plasmid (Invitrogene, USA) under the control of cytomegalovirus promoter and in frame with a 142-bp fragment encoding a modified influenza hemagglutinin cleavable N-terminal signal peptide (MKTIIALSYIFCLVFA) (L Hein et al. 1997 Mol Endocrinol 11: 1266-1277). The construct encoding the chimeric gp82 protein was transiently transfected into Vero cells. To determine the cellular localization of gp82, we analyzed the cells by immunofluorescence followed by confocal laser scanning microscopy. Immunofluorescent labeling of live Vero cells transfected with gp82 construct showed that gp82 was not expressed on the cell surface (not shown). For this reason, we decided to detect gp82 in Vero cells permeabilized with saponin. After 48 hr, cells were fixed in formaldehyde and examined by immunofluorescence followed by confocal laser scanning microscopy. Prior to use, cells were permeabilized with 0.1% saponin and incubated with a monoclonal antibody to gp82, MAb 3F6 (MMG Teixeira & N Yoshida 1986 Mol Biochem Parasitol 18: 271-282), for 1 hr at room temperature, washed three times with phosphate buffered saline and developed with rabbit IgG Cy3-conjugated -Anti Mouse Ig diluted 1:100 (Sigma, USA) for 1hr in the presence of 200 nM DAPI (Molecular Probes, USA) to label DNA-rich structures and either 50 µg/ml 3,3'-dihexyloxacarbocyanine iodide (DiOC6) (Molecular Probes, USA) to label endoplasmic reticulum or 10 µg/ml of wheat germ agglutinin (WGA)-FITC (Sigma, USA) to label distal Golgi complex compartments. The signal peptide from haemagglutinin protein in frame with gp82 allows targeting to endoplasmic reticulum membrane and glycosylation of gp82 protein in transfected cells (MI Ramirez et al. 1998 Mem Inst Oswaldo Cruz 93 Suppl. II: 53-54). Here we show, by immunofluorescence staining of transfected Vero cells with MAb 3F6 (Fig. 2), that gp82 is transported out of the endoplasmic reticulum and Golgi complex and accumulates near the cell surface. When cells were double labeled with MAb 3F6 and WGA-FITC or MAb 3F6 and DiOC6, a poor correlation in the distribution of the these markers was observed (not shown). The gp82 synthesized by the mammalian cells failed to be transported to the cell surface, as also judged by the absence of this protein in the culture medium or on the plasma membrane. Our data also suggested that gp82 expressed in transfected Vero cells lacked GPI modifications. We searched for a putative cleavage/attachment site for GPI anchor addition (w site) at the carboxy-terminal region of gp82 encoded by clone J18 using the w , w +2 rule proposed by S Udenfriend and K Kodukula (1995 Methods Enzymol 250: 571-582). Probabilities of putative w site in the cDNA-deduced carboxy-terminal region were calculated using the hierarchical values of amino acids substituents at sites w , w +1 and w +2 of several GPI-anchored membrane proteins (Udenfriend & Kodukula loc. cit.). The carboxy-terminal domain of gp82 encoded by clone J18 has a putative cleavage/attachment site located from Asp492 to Ser494 residues (w =Asp, w +1=Glyc, w +2=Ser), separated from the hydrophobic C-terminal sequence by a seven residue spacer (Fig. 1). We also analyzed the C-terminal domain of another member of gp82 family encoded by a cDNA clone (R31, GenBank AF128843) isolated and characterized in our laboratory (RC Ruiz 1998 Trypanosoma cruzi: Associação entre Infectividade e Expressão de Glicoproteínas de Superfície com Atividade Sinalizadora de Cálcio, PhD Thesis, Unifesp, 107 pp). The C-terminal domain of the gp82 proteins is highly conserved and it was possible to identify the same w site (w =Asp, w +1=Glyc, w +2=Ser) in both proteins (Fig. 1). The w site (w =Asp, w +1=Glyc, w +2=Ser) of gp82 differs from that found in GPI linked proteins of mammalian cells in which serine is at site w and alanine or glycine is found at site w +2 (L Gerber et al. 1992 J Biol Chem 267: 12168-12173, Udenfriend & Kodukula loc. cit.). It is noteworthy that the w site of gp82 is identical to that of VSG (variant surface glycoprotein) from T. brucei which was experimentally determined (Moran & Caras loc. cit., Udenfriend & Kodukula 1995 loc. cit.). We extended this analysis to other T. cruzi GPI-linked membrane proteins. The predicted cleavage/attachment site for addition of GPI anchor of known T. cruzi GPI-linked proteins is shown in Table. All T. cruzi GPI-anchored proteins have aspartic acid at position w , most of them (20 out of 26 proteins examined) have serine at position w +2 whereas some have glycine or threonine at that site. Glycine or serine is found at position w +1. Once again, many T. cruzi proteins have w site identical to those of T. brucei VSG (Table). Previous work has demonstrated that the w site of VSG (w =Asp, w +1=Glyc, w +2=Ser) functions poorly for GPI processing in mammalian cells (Moran & Caras loc. cit.). Replacement of aspartic acid at site w by serine, or of serine at site w +2 by alanine, converts the inactive VSG GPI signal to viable signal for mammalian cells (Moran & Caras loc. cit.). The presence of aspartic acid and serine at sites w and w +2 in the majority of GPI-linked proteins of trypanosomes suggests that transamidase of these organisms are different from that of mammalian cells. We could speculate that the binding pocket of mammalian transamidase is not large enough to accomodate two slightly larger residues as aspartic acid and serine at sites w and w +2 present in trypanosome GPI signal. Our findings indicate that T. cruzi membrane proteins have unique signal requirements for GPI attachment and the differences found between the trypanosomes and mammalian host cells could be useful for the development of specific antiparasite drugs. Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99097a.jpg] [oc99097c.jpg] [oc99097b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}