 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 94(5), Sep./Oct. 1999 pp 591-596 Bloodmeal Microfilariae Density and the Uptake and Establishment of Wuchereria bancrofti Infections in Culex quinquefasciatus and Aedes aegypti Cleide MR Albuquerque/+, Vânia MS Cavalcanti*, Maria Alice V Melo, Paulo Verçosa, Lêda N Regis*, Hilary Hurd** Departamento de Zoologia, Centro de Ciências Biológicas, Universidade
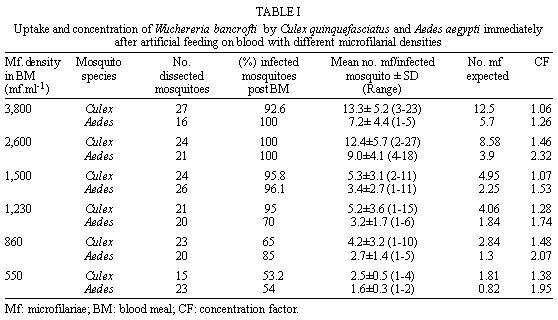
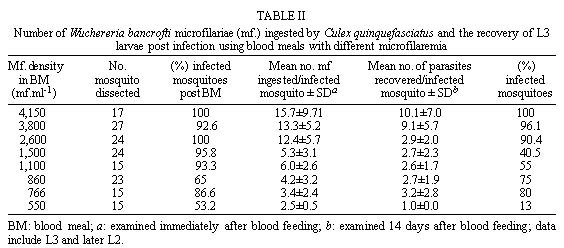
Federal de Pernambuco, Av. Moraes Rego 1235, 50670-420 Recife, PE, Brasil + Corresponding author. Fax: +55-81-271.8359. E-mail: Cleide@npd.ufpe.brReceived 24 November 1998 Code Number:OC99109 The relationship between ingestion of microfilariae (mf), production of infective larvae (L3) and mf density in human blood has been suggested as an important determinant in the transmission dynamics of lymphatic filariasis. Here we assess the role of these factors in determining the competence of a natural vector Culex quinquefasciatus and a non vector Aedes aegypti to transmit Wuchereria bancrofti. Mosquitoes were infected via a membrane feeding procedure. Both mosquito species ingested more than the expected number of microfilariae (concentrating factor was 1.28 and 1.81 for Cx. quinquefasciatus and Ae. aegypti, respectively) but Cx. quinquefasciatus ingested around twice as many mf as Ae. aegypti because its larger blood meal size. Ae. aegypti showed a faster mf migration capacity compared to Cx. quinquefasciatus but did not allow parasite maturation under our experimental conditions. Similar proportions of melanized parasites were observed in Ae. aegypti (2.4%) and Cx. quinquefasciatus (2.1%). However, no relationship between rate of infection and melanization was observed. We conclude that in these conditions physiological factors governing parasite development in the thorax may be more important in limiting vectorial competence than the density of mf ingested. Key words: Culex quinquefasciatus - Aedes aegypti - Wuchereria bancrofti - vectorial capacity Lymphatic filariasis is one of the most important human diseases transmitted by mosquitoes. It affects approximately 120 million people worldwide (Otensen & Ramachandran 1995). Understanding of vector/parasite interactions is essential for rational development of filariasis control measures. This is particularly important, considering that vectorial competence, i.e. the ability of mosquitoes to ingest mf and to promote their maturation until the infective stage and the rate of mosquito survival until parasite maturation (Failloux et al. 1995), seem to differ according to geographic mosquito strain (Wharton 1960, Crans 1973, McGreevy et al. 1982). Variation in the density of mf in the blood and parasite behaviour also influence vector/parasite relationships (Tabachnick et al. 1985, Southgate & Bryan 1992, Failloux et al. 1995). Recife, a Brazilian coastal city (on 8o4'3" south latitude), with approximately 1.4 million people, is an endemic area of Wuchereria bancrofti, transmitted by Culex quinquefasciatus. In 1952, Azevedo and Dobbin registered lymphatic filariasis in a local district with microfilaraemia rate of about 9.7%. A recent survey, of several areas, has shown that W. bancrofti infection now has prevalence rates ranging from 0.64 to 14.95% (Maciel et al. 1996). In addition to increasing incidence of lymphatic filariasis in some areas, the number of breeding places for Cx. quinquefasciatus is rising due to uncontrolled urbanization (Oliveira 1996). Despite these facts, we have a poor understanding of the interactions that occur between the parasite and its vector in Recife. Ae. aegypti is also abundant in Recife, but it is not regarded as a natural vector of W. bancrofti, however understanding of the relationship mosquito/parasite is relevant to studies concerned to immune mechanism of refractory species. It has been suggested that the relationship between the density of microfilariae (mf) in human blood, the ingestion of mf and production of L3 larvae is an important determinant of transmission dynamics of lymphatic filariasis (Brito et al. 1997, Calheiros et al. 1998). Ingestion may depend upon feeding behaviour and mechanics, whereas, difference in vector physiology will contribute to the success or failure of parasite maturation to L3. Here we report upon a study designed to investigate the relative role of these factors in the development of W. bancrofti in the susceptible Cx. quinquefasciatus and refractory Ae. aegypti and describe our initial observations on the immune response to mf in these two mosquitoes. MATERIALS AND METHODS Mosquitoes - Laboratory bred Cx. quinque-fasciatus (Recife strain) and Ae. aegypti (Recife strain) were used throughout this study. Colonies were obtained from field collected mosquitoes in endemic areas in Recife. Cx. quinquefasciatus has been maintained in the laboratory for four years and Ae. aegypti for approximately one year. Adults were maintained in cages (30x30x30 cm) at room temperature (27±3oC) with 85±10% relative humidity, and fed with 10% (w/v) sucrose solution ad libitum. Infection via artificial blood feeding - Experimental feeding was carried out using four to seven day old female mosquitoes, the sugar meal having been withdrawn 12 hr beforehand. Blood was obtained from selected human lymphatic filariasis carriers, who participated in a survey carried out by Centro de Pesquisas Aggeu Magalhães-Fiocruz, Recife, after written informed consent. Volunteers received no previous anti-microfilarial treatment and possessed different densities of circulating mf. At the end of the work, donors were treated with diethylcarbamazine according to therapeutic recommendation of the World Health Organization (WHO 1987). Venous blood was collected between 23-24 hr in vacuntainers containing EDTA (1 mg. ml.-1) as an anticoagulant. The blood was kept at 4oC until the next morning. Microfilariae density was estimated by counting the number in 60 mm3 (3x20 mm3 aliquots) of blood and expressed as mf/ml. Number of parasites in the blood ranging between 550-860 mf/ml-1 was considered low density (LD), between 1,230-1,500 mf/ml-1 as medium density (MD) and between 2,600-4,150 mf/ml-1 as high density (HD). Groups of 300-400 mosquitoes per cage were allowed to feed for 1 hr on the infected blood, heated to 37oC, using glass feeders covered with cow gut membranes (Rutledge et al. 1964). To ensure microfilariae homogeneity the blood was kept agitated using a stirrer at 1 red/sec.-1 Only females which were fully engorged were used in the experiments. Dissection - Approximately 20 females of each mosquito species were dissected immediately after feeding, to determine the number of microfilariae ingested. Similar numbers of engorged mosquitoes were dissected at 4 hr, 24 hr, 72 hr, 7 and 14 days following the blood meal, to assess migration and development of parasites. The head, thorax and abdomen of mosquitoes were examined for second and third filariae larval stages. Estimation of blood meal size and microfilariae concentration - Blood meal volume ingested by mosquitoes was estimated by weighing 120 of each mosquito species before and after the blood meal. The weight difference was attributed to ingested blood. Conversion of the average weight gain to volume was achieved by dividing the weight gain by the specific gravity of blood (1.05 mg.mm3-1). The expected uptake of mf was calculated based on Bryan and Southgate (1988). Concentration factor was estimated by dividing the observed mf intake by the expected mf ingested. Cx. quinque-fasciatus and Ae. aegypti were dissected to determine the relationships between mf density in the blood of the donor and subsequent uptake of mf, migration, development and mosquito concentrating effect. Nine experiments were performed using Cx. quinquefasciatus and six trials were carried out simultaneously with Ae. aegypti. Statistical analysis - Linear regression analysis was used to compare mf blood density and mean number of parasites ingestes per mosquito. Data were normalized after log10 transformation, with zero values being excluded from the samples. Similar analysis was performed to relate the density of mf in blood to the number of larvae recorvered 14 days after infection. Differences in mf ingestion between Cx. quinquefasciatus and Ae. aegypti at the same parasite densities were tested using Student's t-test. Variations in the proportion of Cx. quinquefasciatus and Ae. aegypti females infected were compared by ANOVA. RESULTS Weight of blood meal - Mean weights of blood meals were 3.5 mg and 1.6 mg per insect, for Cx. quinquefasciatus and Ae. aegypti, respectively. These weights converted to mean volumes of 3.3 mm3 and 1.5 mm3 for the two species. Relationship between mf density in the blood and uptake of mf by mosquitoes - The mean number of mf ingested by both mosquito species at different mf densities is presented in table 1. Additional trials performed with only Cx. quinque-fasciatus are shown in table 2. Blood density of mf was positively correlated with mean number of parasites ingested by both species of mosquitoes. Considering all nine experiments for Cx. quinquefasciatus and six, for Ae. aegypti, values obtained for the correlation of blood mf density and mean number of mf ingested per mosquito were: Cx. quinquefasciatus r = 0.988; t = 15.37; P = 0.000; the regression line is represented by the equation y = -2.05+ 0.898x; and Ae. aegypti r = 0.948; t = 6.69; P = 0.001 and y = -2.40 + 0.935x. In both species, the mean number of mf ingested exceeded the expected number, based on the size of the blood meal and the mf density in the blood. Thus, Ae. aegypti and Cx. quinquefasciatus were able to concentrated W. bancrofti mf; although Ae. aegypti was more efficient than Cx. quinque-fasciatus (1.81 and 1.28 times respectively). Differences in blood meal size tended to be compensated by differences in ability to concentrate parasite, however, Cx. quinquefasciatus (larger blood meal) always ingested more mf than Ae. aegypti. The ability to concentrate mf increased as mf densities decreased, in both mosquito species ( table 1). The incidence of infection was similar when Cx. quinquefasciatus and Ae. aegypti were compared after feeding on same microfilariae density in the blood ( table 1). This was observed in all trials except for those with a mf density of 1,230 and 860 mf/ml-1. At this microfilariae densities a significant difference was observed between Cx. quinquefasciatus and Ae. aegypti (ANOVA F=1.93 P<0.05). Comparison of parasite migration in each species - Migration of mf from the midgut to the thorax seems to be unaffected by the density of mf in the blood ( table 3). The rate of mf migration was faster in Ae. aegypti than in Cx. quinquefasciatus. At 4 hr approximately 40% more parasites were observed in the thorax of Ae. aegypti. Migration continued until 72 hr when the majority of the parasites (80-100%) were observed in the thorax in both mosquito species ( table 3). In Cx. quinquefasciatus, by day 14 approximately, 17.1% of the L3 (range 5.6-35%) had migrated to the head, while the majority (73.8%) had migrated within 21 days. Relationship between mf uptake and number of larvae recovered from Cx. quinquefasciatus, 14 days following an infective blood meal - Immediately after feeding, the proportions of infected Cx. quinquefasciatus females ranged from 53.2 to 100% (eight experiments) ( table 2). After 14 days, the proportion of infected females varied between 13 and 100% ( table 2). Infective larvae could occasionally be observed as early as 13 days post infection. Microfilarial densities of less than 1,500 mf/ml-1 significantly affected the proportion of infected female (P<0.001). The mean number of parasites ingested varied between 2.5±0.5 and 15.7±9.7, mf/infected female at day 0. A proportion of the mf ingested was lost in the mosquito host during development to the infective stages so that, by day 14, L2 and L3 parasite densities ranged between 1±0 and 10.1±7/infected female. No relationship was found between mf density in the blood and the number of mf surviving to maturity. The regression line is represented by the equation y = -0.00003133 - 0.763 x, r = 0.0 (P<0.05). No infective larvae were found in Ae. aegypti, 14 days after the infective blood meal. However, in the same period undeveloped parasites were observed in the thorax of approximately 85% of females fed with blood containing 1,500 and 2,600 mf/ml-1. Throughout the study, a small number of melanized larvae were recovered from both mosquito species assessed until three days post infection. From a total of 997 parasites recovered in Ae. aegypti 2.4% showed melanization. Similar proportions of melanized parasites were also recovered in Cx. quinquefasciatus (2.1%). Melanization was observed as early as 4 hr following an infective blood meal in the abdomen. An intracellular melanization response to first-stage larvae in the thoracic muscle fibres was also observed between 24-72 hr after the infective blood meal. DISCUSSION It has long been recognized that intrinsic difference in the dynamic relationship between W. bancrofti and its vectors necessitates the performance of detailed studies in each endemic area (Samarawickrema & Laurence 1978, McGreevy et al. 1982, Lowichich & Lowrie 1988, Brito et al. 1997). This study provides the first date of this kind for Recife. Our data confirm the positive correlation between microfilariae density in the donor blood and the number of microfilariae ingested by mosquitoes that has been described by others (Obiamiwe 1977, McGreevy et al. 1982, Bryan & Southgate 1988, Janousek & Lowrie 1989, Jayasekera et al. 1991). McGreevy et al. (1982) suggested that for a variety of Culex, Aedes and Anopheles species, variations in mf uptake reflected the mean volume of blood ingested. In our study this also appear to be true. Cx. quinquefasciatus was less able than Ae. aegypti to concentrate mf from the host blood, yet always ingested more microfilariae probably due the ability to take up approximately twice as much blood (see table 1). Although other factors may influence microfilarial uptake, it seems likely that blood volume ingested is the major difference between the two species in this study. The relationship between blood microfilaria density and the capacity of vector to concentrate W. bancrofti has been described in several reports. In the majority of these studies, as in the present one, concentrating capacity decreased as mf densities increased (Samarawickema & Laurence 1978, Bryan & Soulthgate 1988, Lowichik & Lowrie 1988, Jayasekera et al. 1991). An increase (³ 2x) of mf in the mosquito compared with the number expected on the bases of mf density in the blood is common. Gad et al. (1996) reported a two fold concentrating effect for Cx. pipiens and Ae. caspius based on volumes of 2.8 mm3 and 2.3 mm3, respectively. Different concentrating capacities of Cx. quinquefasciatus (Haitian strain) and Ae. aegypti (Liverpool strain) were described by Lowichik and Lowrie (1988). However, they observed that Cx. quinquefasciatus was more efficient in concentrating mf (3.2 - fold factor) than Ae. aegypti (2.6 - fold factor). The opposite to those obtained in the present work (1.28 for Cx. quinquefasciatus and 1.81 for Ae. aegypti). Our data tend to indicate that, at a range of infection doses, between 550 and 2,600 mf/ml-1, the number of parasites that develop successfully in Cx. quinquefasciatus is limited, and independent of microfilariae density in the blood meal (1 to 3.2 per mosquito). Brito et al. (1997) reporting on a study in Maceió e Belo Horizonte found a similar range of L3/mosquito (1.62 and 2.62) after feeding on donor with 450 and 700 mf/ml-1, respectively. Southgate and Bryan (1992) suggested some limitation to development for filarial infections in Cx. quinquefasciatus. However, when higher infective dose were used in our study this limitation appear to brake down giving rise to much higher parasite development rates as shown in table 2. At concentrations of mf ranging from 2,600 ml-1 to 3,800 ml-1 in the blood, we found that the number of larvae subsequently recovered increases from 2.9 to 9.1 per mosquito. This phenomenon of facilitation, where numbers of developing larvae increases disproportionately to infective dose, is currently under discussion (Wada et al. 1995). Janousek and Lowrie (1989) found that 50% of the Wuchereria ingested by Cx. quinquefasciatus (Haitian strain) reached the muscles after 18 hr post blood meal. In the present study, migration from the blood meal to the thorax muscle occurred more quickly in Ae. aegypti (approximately 88.8% 24 hr post feeding) than Cx. quinquefasciatus (36.6%). Migration continued until 72 hr with majority of larvae found in the thorax, in both species. From these data we conclude that Ae. aegypti can successfully be infected with W. bancrofti, it has a higher concentrating factor and showed a faster mf migration capacity compared to Cx. quinquefasciatus. However, Ae. aegypti did not allow parasite maturation under our experimental conditions. Differences on parasite development between the two species appear to occur once the parasite are in the thorax rather than when mf are passing through the gut or haemolymph, where the vector defense mechanisms are known to be stimulated. If nutrient competition between parasite and vector is limiting parasite development it is conceivable that the larger blood meal taken by Cx. quinquefasciatus contributes to its vectorial competence. Multiple blood meals have been shown to enhance filarial development (Ham & Gale 1984, Travi & Orhiel 1987). Our study used artificial membrane feeding in which only one blood meal was offered to the mosquitoes at the time of infection. Provision of additional blood feeds may have enhanced W. bancrofti development in Ae. aegypti. However this species is not a natural vector of filariasis, although additional meals will be taken in the field. Lowichik and Lowrie (1988) observed that the laboratory selected Liverpool strain of Ae. aegypti is able to support higher levels of development of W. bancrofti than seen in Cx. quinquefasciatus. It is probable that differences in vectorial competence are due to variation in the genetic response of the mosquitoes to infection, although how these differences operate is unknown at present. Small proportions of melanized parasites were observed in both mosquito species, suggesting defence mechanisms in both susceptible and refractory strains. However, other physiological processes must be involved in limiting parasite development since melanization alone can not explain the underdevelopment of these parasites in Ae. aegypti. Furthermore, no relationship between proportion of mf ingested and melanization was observed in our work as described in a refractory strain of An. quadrimaculatus infected with Brugia malayi (Nayar et al. 1992). In summary, studies on the relationship between Cx. quinquefasciatus and W. bancrofti from Recife, resulted in a vector competence of 53% (based on Janousek & Lowrie 1989). This vector competence is among the highest reported in the literature and may account, at least, in part, for high mf rate in Recife (14.7%) (Maciel et al. 1996). ACKNOWLEDGEMENTS To Dr André Furtado for valuable discussion; to Mrs EHM Rocha for technical assistance. This work was supported by the Wellcome Trust and CNPq. REFERENCES
Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99109a.jpg] [oc99109b.jpg] [oc99109c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}