 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
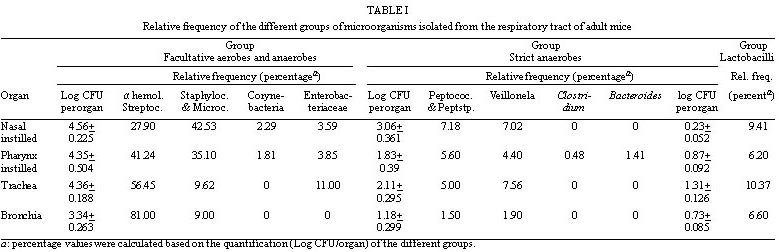
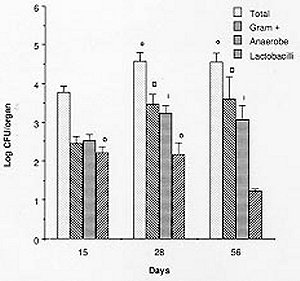
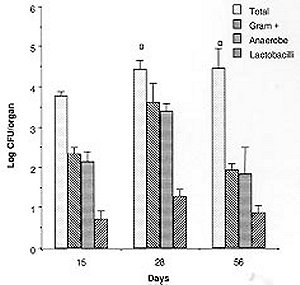
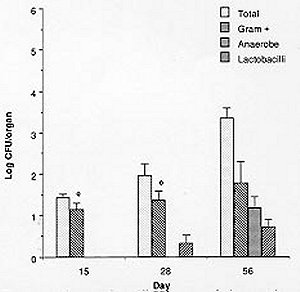
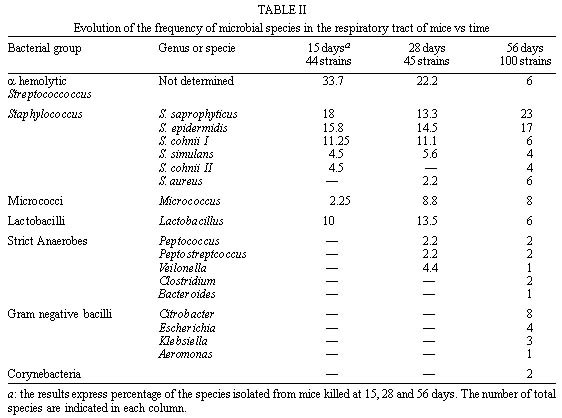
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 94(5) Sep./Oct. 1999, pp: 701-707 Microbial Flora Variations in the Respiratory Tract of Mice Rosa Cangemi de Gutierrez, Olga Miguel de Nader, Aida Pesce de Ruiz Holgado*, María Elena Nader-Macias*/+ Instituto de Microbiología, Facultad de Bioquímica, Química y Farmacia, Universidad Nacional de Tucumán, Tucumán, Argentina *Cerela (Centro de Referencia para Lactobacilos), Conicet, Chacabuco 145, 4000 Tucumán, Argentina + Corresponding author. Fax: +381- 431-1720. E-mail: mmacias@CERELA.org.ar.Received 8 September 1998 Code Number: OC99131 A stable microbial system in the respiratory tract acts as an important defense mechanism against pathogenic microorganisms. Perturbations in this system may allow pathogens to establish. In an ecological environment such as the respiratory tract, there are many diverse factors that play a role in the establishment of the indigenous flora. In the present work we studied the normal microbial flora of different areas of the respiratory tract of mice and their evolution from the time the mice were born. Our interest was to know which were the dominant groups of microorganisms in each area, which were the first capable of colonizing and which dominated over time to be used as probiotic microorganisms. Our results show that Gram negative facultatively anaerobic bacilli and strict anaerobic microorganisms were the last ones to appear in the bronchia, while aerobic and Gram positive cocci were present in all the areas of the respiratory tract. The number of facultative aerobes and strict anaerobes were similar in the nasal passage, pharynx instilled and trachea, but lower in bronchia. The dominant species were Streptococcus viridans and Staphylococcus saprophyticcus, followed by S. epidermidis, Lactobacilli and S. cohnii I which were present on every studied days but at different proportions. This paper is the first part of a research topic investigating the protective effect of the indigenous flora against pathogens using the mice as an experimental model. Key words: indigenous microflora - colonization - respiratory tract - probiotics The indigenous microflora of human beings and animals is a remarkably stable ecosystem. Some variations exist in the flora composition and in the number of different organisms, but within a given individual, the flora remains relatively constant with time (Lidbeck et al. 1987). These microbial flora are in ecological balance, protecting the host from pathogens. Different mechanisms have been proposed to explain this effect including the production of inhibitory substances as bacteriocins (Jack et al. 1995), organic acid, hydrogen peroxide (Klebanoff et al. 1991), competitive exclusion (Chan et al. 1984), steric hindrance (Chan et al. 1985), competition for nutrients (Freter et al. 1983) or immune system stimulation (Perdigón et al. 1988). The ecological balance can be disrupted by many factors: some of them host dependent (immunological or oncological states), extrinsic (antibiotic or hormone treatments) or microorganism-dependent ones. Suppression of the indigenous microorganisms often permits overgrowth by potential pathogens resulting in many microbial pathologies (Lidbeck et al. 1987). The human oropharynx is ordinarily colonized by aerobic and microaerophilic bacteria, along with anaerobes (Niederman 1990). The mucosa of the nasopharynx, trachea and the major bronchia are also colonized with aerobic and anaerobic bacteria. This epithelial surface coexists with the microbial flora and is not overgrown by it. Moreover, the physiological function of the mucosa - including a protective barrier, mucociliary clearance and humidification, and warnings of respired air - are not impeded (Reynolds 1987). The respiratory tract infections are among the most frequent health problems in underdeveloped countries (OMS 1994), with many having bacterial or viral origins. Infections of the lower respiratory tract affect 4 to 5 million people in America, with pneumonia representing the leading cause of death due to infection. In the last years, there has been an increased tendency in the modern medicine to use more preventive rather than curative treatments, together with natural products. Some of these products are the "probiotics" that have proven to be beneficial for both human and animal health. The term "probiotics" defined by Havenaar et al. (1992) are viable cultures of one or several microorganisms that, when administered to human or animals, produce a beneficial effect on the host, by fulfilling the role of the normal endogenous microflora. Preparations containing lactobacilli have been administered to humans to prevent or cure diarrhea (González et al. 1990), as well as bacterial and mycotic vaginitis (Redondo-López et al. 1990), in experimental models using mice (Nader de Macías et al. 1996). Subsequently, the concept of re-establishing the ecology of those tracts randomly colonized by "normal" or "indigenous" microflora to contribute to the maintenance of this balance has lead many researchers to study the protective role of the indigenous microflora. This is also the motivation of our group: to study the indigenous flora of the respiratory tract of mice and assess their protective effect. In our research area this type of investigation is focused in aerosolized products to be used in the respiratory tract. We were unable to find in the present literature a report refering to this topic using mice as experimental model. The present paper shows our results with reference to (a) the kinetics of colonization of the most frequent groups and species of microorganisms in the respiratory tract of mice; (b) their levels in young and adult mice, together with (c) the most frequent species isolated from different areas of the respiratory tract by using mice as experimental model. Our next step will be to test and select those effective and non pathogenic species to protect the mice against pathogens. MATERIALS AND METHODS Animals - BALB/C male mice from the stock breeding colony held in our laboratory were used throughout this study. The animals were housed in plastic cages and fed ad libitum under constant environmental conditions. Each experiment was carried out with groups of 15 to 30 mice. They were sacrificed on days 15, 28 and 56 after birth. Culture media and growth condition - Samples were plated in duplicate on different enriched media: Blood Agar (Columbia Agar with 5% blood, and Columbia Agar with 5% heated blood 15 min at 70°C), Brain Heart Infusion (BHI) supplemented with 5mg/ml hemin, 0.005% cistein chlorhydrate, 0.5% yeast extract, 100 mg/ml Kanamicin and 1 mg/ml vitamin K for anaerobes (Sutter et al. 1978), and selective media including Lactobacillus Selection Media (LBS) (De Man et al. 1960) and Manitol Salt Agar. The plates were incubated at 37°C for 24, 48 or 72 hr, in aerobic and anaerobic conditions. The microorganisms were identified according to standard methodologies using biochemical tests (Bannatyne et al. 1979). Lactobacilli were identified by the methods described in Bergey's Manual of Determinative Bacteriology (Kandler & Weiis 1986), as well as standard techniques used in our laboratory, and API 50 CHL (Biomerieux-France). Bacterial counts in tissue homogenates - Mice were sacrificed by cervical dislocation. Nasal and pharynx instilled were obtained by washing each opened cavity with 1 ml of peptone water, and later scrapping them softly with sterile cotton tips. Trachea, bronchia and lungs were aseptically removed, rinsed and homogenized in 1 ml peptone water with a Teflon pestle. Dilutions were made and the samples plated in duplicate on the different culture media. Plates were incubated for 48-72 hr aerobically or anaerobically. Counts of viable cells were made for each organ. Isolation and identification of the microorganisms were performed as described before. Results were expressed as the mean ± standard deviation (SD) of the data obtained from all the animals. Statistical analysis - The results show the mean of the data obtained from 15 to 30 mice ± the SD. The experiments were performed at least twice. Analyses of the data variance were used to study the importance of the differences between the data (test f). RESULTS Quantification of microorganisms in the respiratory tract - The levels of the different groups of microorganisms studied (expressed as Log CFU/organ) in the various areas of the respiratory tract of adult mice are shown in Table I. The number of microorganisms obtained in the bacterial groups or genus was used to calculate the relative frequency of each of them and are shown as percentage values. The quantification of microorganism showed similar values in the total facultative aerobes and anaerobes in nasal, pharynx instillated and trachea showing no statistical different result (p>0.01). In bronchia their levels were one log unit lower.The strict anaerobes showed similar value (p>0.01) in the pharynx and trachea, being lower in bronchial samples and higher (p>0.01) in nasal instilled. Lactobacilli were present in low numbers in the four areas studied. Based on their distribution we can observe that (1) the alpha hemolytic streptococci increased in frequency as samples were collected further down the respiratory tract, and represented the dominant group of microorganisms; (2) by contrast, Staphylococcus and Micrococcus decreased in frequency from nasal instilled to bronchia, being the sub-dominant groups; (3) Corynebacteria were present only in nasal instilled and pharynx in very low numbers; (4) Gram negative bacilli levels increased until the trachea and were not present in bronchial samples; (5) the strict anaerobes showed different levels in the areas, decreasing significantly in bronchia. Clostridium and Bacteroides were only present in pharynx; (6) the low levels of Lactobacillus in the four areas were represented by isolates of L. brevis, L. fermentum ss. cellobiosus and L. plantarum. Kinetics of colonization of microorganisms vs time Microorganisms in nasal instilled - Fig. 1 shows that the number of total aerobes, Gram positive cocci and strict anaerobes follow the same type of kinetics, increasing in number up to the 28th day, and maintaining their levels in the adult mice. The amount of lactobacilli in levels of log CFU/organ=2, decreased on the 56th day. The paired symbols in the graph show no statistically significant differences (p> 0.01). Microorganisms from pharynx - Samples of pharynx, there was also a slight increase in the number of total aerobes, while the Gram positive cocci, strict anaerobes and lactobacilli were higher on the 28th day, but decreased on the 56th day to almost the same level recorded on day 15 (Fig. 2). Microorganisms from trachea - In trachea, the total aerobes and strict anaerobes decreased slightly on day 28, but increased their levels again by day 56. The Gram positive cocci and the lactobacilli increased on day 28, and maintained their levels on day 56 (Fig. 3). Microorganisms from bronchia - The most important characteristic was the low number of total aerobes on all the days studied. The Gram positive cocci were in the same level. The lactobacilli appeared in very low numbers on day 28, increasing slightly on day 56. The strict anaerobic microorganisms were present only on day 56, also at low levels. This last group appeared as part of the adult microflora of the mice (Fig. 4). Species or genus of microorganisms isolated at different times - The genus or species of microorganisms isolated in all the areas of the respiratory tract of mice at 15, 28 and 56 days are shown in Table II. Considering all the strains together, we can observe the percentage of different genus or strains isolated in each sampling-time: 44, 45 and 100 different isolates from days 15, 28 and 56 respectively. The studies at the species level were performed only in those groups of microorganisms with potential application as probiotics, but not in the other groups. The results obtained were as follows: (1) the predominant number of strains isolated within 15 and 28 days old mice were the a hemolytic Streptococcus, which decreased on day 56; (2) in the Staphylococcus group, the percentage of S. saprophyticcus strains showed a slight tendency to increase its number while S. epidermidis maintained very similar levels all time points; (3) the frequency of Micrococcus strains increased slightly between 15 and 28 days, maintaining the ratio up to 56 days; (4) lactobacilli were present in the three age groups of mice with very limited number of different strains; (5) the Gram negative bacilli (16 strains) and Corynebacteria (2 strains) were only present in the adult mice at very low percentages; (6) some strict anaerobe species appeared on day 28, while the others were only recorded in the adult mice (8 strains), showing a very low frequency.DISCUSSION Soon after birth the open cavities of man and animals are colonized by a wide variety of microorganisms. They become established in the oropharynx, intestinal and vaginal tracts, constituting the "indigenous" microflora. This normal indigenous flora can act competitively excluding pathogens, thus inhibiting their colonization and consequently preventing infection as stated by many authors (Herthelius et al. 1989). However, this is true only for certain components of this indigenous flora, and for certain pathogens. Based on the host-specificity that characterizes the indigenous flora (Savage & Kotarsky 1979), we used mice as an experimental model to study the protective effect of the microorganisms that are part of this microflora. In this paper we report the evolution of the different groups of bacteria with regards to their quantitative dominance over two months; from the time the mice were born and considering the mice as adult at two months old. This was performed to evaluate which were the first groups of microorganisms capable of colonizing the different areas. There is a lot of work published on the application of probiotics in the gastrointestinal tract (Havenaar et al. 1992). Also, the requirements for the strains to be used in this area supported with experimental models developed to explain that type of research (Perdigón et al. 1988). For example some studies have been performed examing the digestive tract and feces of man and animals from birth (Ducluzeau 1989), showing which were the dominant and subdominant groups, and the mechanisms involved in their regulation (Freter et al. 1983). With regards to the respiratory tract, there have been studies reporting which are the microorganisms present in the respiratory tract of man in order to avoid contamination by them when trying to isolate pathogenic bacteria and for the interpretation of cultural results (Bartlett et al. 1987). It has been reported that alpha-hemolytic and non-hemolytic streptococci, Haemophillus sp., Corynebacteria, Staphylococci, Micrococci and anaerobic bacteria are present. Our results showed that the dominant and subdominant groups were alpha hemolytic streptococci, Staphylococcus and Micrococcus groups in mice. We suggest that the presence of these groups is regulated by the local, ecological and physiological conditions in each of the areas, which differ from each other. Some of these situations are non-specific, because of the amount and type of different available nutrients, pH, redox potential and/or resistance to local antibacterial substances. Some are specific, including the nature of the adhesion promoting factors for each of the bacterial groups, plus the specific receptors either in the epithelial cells or mucus gel. S. epidermidis and diphtheroids in man as described by Salyers and Whitt (1994) often colonize the anterior nares. A paper reporting the role of the normal throat flora in prevention and colonization by Group A streptococci was published by Crowe et al. (1973), where they suggested that certain constituents of the normal flora such as non hemolytic streptococci, viridans streptococci and Neisseria species are implicated in colonization resistance of the respiratory tract to both group A streptococci and Gram-negative bacilli. There is no report of the evolution of the species in man or animals, or information detailing which are the first species colonizing, as we have shown in the present paper. Our most important finding was that the anaerobe group was the last one colonizing the bronchia, together with lactobacilli which appeared before. The anaerobic flora in man is around 103-104 CFU in nasal instilled (Tramont 1990). With regards to the species isolated, they are Peptococcus, Peptostreptococcus, Veillonella, Clostridium and Bacteroides, the first three in 28 days old mice, and all of them in 56 days old mice. We isolated Gram negative bacilli (Citrobacter, E. coli, Klebsiella pneumoniae, and Aeromonas) as part of the normal flora of nasal and pharynx instilled in higher amounts than in tracheal samples. Niederman (1990) discussed the pathogenicity and clinical consequences of Gram negative colonization of the respiratory tract referring to colonization as the microorganisms exerting their pathogenicity and virulence. The normal upper and lower human tract are not colonized by enteric Gram negative bacteria, which can be harbored when serious illness develops, as described the same author, who stated that their colonization increased when severity of the illness is greater. The influence of topical antimicrobial prophylactics in colonization of oropharynx and trachea in patients in Intensive Care Units has also been studied (Bontem et al. 1994). The presence of Gram negative bacilli in chronically ventilated patients, Pseudomonas and members of the family Enterobacteriaceae in trachea and oropharynx (Palmer et al. 1995) was reported. Our concept of colonization is not related with pathogenicity, instead it is related to that defined by Freter (1992) for the normal microflora of the gastrointestinal tract. He stated that colonization describes a bacterial population that is stable in size over time, without the need for periodic re-introduction of the bacteria. The evolution of the respiratory tract microflora evolve from the birth depending on which are the first microorganisms in contact with the new born animal. Later, as they are exposed to a complete spectra of microorganisms, the predominant ones are those able to adapt or to colonize certain areas of the mucosal tree. Then, the microflora of the animals grown in the same environment are very closely related to the same animals, and stable under the established conditions. Another factor to be considered in the stablishment or presence of the microorganisms as indigenous flora of adult individuals, referred to mice, is the coprophagy characteristic of this group of animals, what in some way, can influence the adult microflora equilibrium. Lactobacilli are present in the entire respiratory tract in low levels. There is no report of the presence of lactobacilli as part of the indigenous microflora of the respiratory tract, with the exception at Liedbeck et al. (1987) who reported the impact of L. acidophilus supplements on the human oropharynx and intestinal microflora. They determined the number of lactobacilli in saliva and feces of volunteers fed with L. acidophilus milk. They reported values of lactobacilli between 102 and 103 CFU/ ml of saliva and suggested that the quantification of microorganisms from saliva reflected the number of bacteria in the oropharynx area. The authors did not describe the method of obtention of saliva samples, either with stimulation or not, because while obtened stimulated saliva samples, the number of lactobacilli present in the dental plaque of teeth are extracted. Then, the number of lactobacilli in saliva samples could be incremented by this reason. We were interested in knowing what are the kinetics of colonization of the different groups of microorganism, and we quantified their numbers over two months while mice were growing from their birth. Our results allow us to perform an evaluation of which are the dominant groups, genus and species present in the different areas of the respiratory tract, and how are they modified over the time. All this information could be helpful in the establishment of criteria to select the strains to be used as probiotics using the mouse experimental model. We are testing some other in vitro characteristics of the isolated strains to choose those optimal strains to be used to protect the mice from pathogenic bacteria challenge. ACKNOWLEDGEMENTS To Lic. Elena Bur for her assistance and valuable help in the statistical evaluation of the data. REFERENCES
Copyright 1999 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc99131a.jpg] [oc99131c.jpg] [oc99131b.jpg] [oc99131f.jpg] [oc99131e.jpg] [oc99131d.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}