
|
Indian Journal of Occupational and Environmental Medicine
Medknow Publications
ISSN: 0973-2284 EISSN: 1998-3670
Vol. 9, Num. 2, 2005, pp. 65-70
|
Indian Journal of Occupational and Environmental Medicine, Vol. 9, No. 2, May-August, 2005, pp. 65-70
Original Article
Vitellogenin assay by enzyme-linked immunosorbant assay as a biomarker of endocrine disruptor chemicals pollution
Ebrahimi M
Environmental Hormones Pollutants Research Center,
University of Qom, Qom, Iran
Correspondence Address: Dr. M. Ebrahimi, Environmental Hormones Pollutants Research
Center, University of QOM, QOM, Iran. ebrahimi@qom.ac.ir
Code Number: oe05016
Abstract There is increasing evidence that many xenobiotic chemicals (called as endocrine disruptor chemicals, EDCs) through interfering with the endocrine system, have the capability to induce developmental and reproductive abnormalities in humans and animals. The yolk protein precursor vitellogenin (Vtg) has proved to be a simple and sensitive biomarker for assessing exposure of fish to EDCs, especially the estrogenic compounds. Work is ongoing to develop screening and testing programmes for endocrine disrupting effects of new chemicals, and in the focus of this development are the fish test species common carp ( Cyprinus carpio ). In this study, we have developed quantitative enzyme-linked immunosorbent assays (ELISA) for VTS in common carp. The working range of the ELISA was 11.25 - 2000 ng/ml (75 - 25% specific binding/maximum antibody binding [B/B0]) with a 50% B/B0 intra and interassay variation of 3.9% ( n = 10) and 12.5% ( n = 30), respectively. This ELISA is capable of detecting Vtg as low as 6 ng/ml, and can accurately detect Vtg in even 10 ml of plasma. The ELISA was applied to measurement of Vtg production by male carp ( C. carpio , Cyprinidae) fish exposure to ethynylestradiol. The results showed that the amount of Vtg produced in plasma of exposed fish increased in logarithmic order comparing to the control group and the ELISA described here could be used as an indicator of water pollution to estrogenic pollutants.
Keywords: ELISA, Endocrine disruptor chemicals, Immunoassay, Vitellogenin
Introduction
The last two decades have witnessed growing scientific concerns and public debate over the potential adverse effects that may result from exposure to a group of chemicals that have the potential to alter the normal functioning of the endocrine system in wildlife and humans.[1] According to WHO definition, an endocrine disruptor chemical (EDC) is an exogenous substance or mixture that alters function(s) of the endocrine system and consequently causes adverse health effects in an intact organism, or its progeny or (sub) populations. The EDCs encompass a variety of chemical classes, including natural and synthetic hormones, plant constituents, pesticides, compounds used in the plastics industry and in consumer products, and other industrial by-products and pollutants.[2]
Mammalian studies have focused on the male since it is much easier to determine changes in the quality and quantity of sperm than of eggs, because eggs are few and are retained internally.[3] Fish have a major advantage over mammals in studies of endocrine disruptors in that they produce large numbers of both eggs and sperms and release them to the external environment. Hence, the effects of potential endocrine disruptors can be readily examined in both sexes. An assessment of pollution requires a simple biomarker and Vtg can be the easiest choice.
In very simple terms, vitellogenesis involves ovarian estradiol stimulating
the liver to produce Vtg, which in turn is incorporated into the yolk
of developing oocytes. Estrogenic xenobiotics can also act on the hepatic
receptors to induce synthesis of Vtg.[4] Both
male and female fish, as well as immature juveniles have hepatic estrogen
receptors, but only the livers of female fish will normally be exposed
to estrogens.[5] Production
of Vtg by males and juveniles, or nonvitellogenic females can therefore
provide a bioindicator of exposure to environmental estrogens.[6]
A number of studies have used Vtg to demonstrate the presence of estrogenic
chemicals in the aquatic ecosystem. Trout exposed to sewage effluent
in English rivers had up to 100 000-fold increase in plasma Vtg compared
to fish upstream of the effluent.[7] A
follow-up study in which caged trout were held in the River Aire, which
is polluted by detergents from textile mills, showed Vtg levels of
more than 20 mg/ml 5 km downstream of the effluent.[8] Elevated
plasma Vtg was also found in eels below the Paris outfall.[9] Because
eels may spend up to 20 years in the river before their spawning, migration-consequent
reproductive disruption may account for some of the decrease in the
European eel population.[9] The
flounder, as a bottom-dwelling estuarine species, also provides a useful
biomonitor of effluents in the rivers, which may potentially affect
marine fish stocks.[10] In
another study, male flounder in a number of British rivers had elevated
plasma Vtg and, in the River Mersey, the concentrations were around
100 mg/ml (ten million times higher than controls).[11]
During the last 15 years, there have been a number of publications
in which enzyme-linked immunosorbent assay (ELISA) using microtiter
plates have been described as an alternative to radioimmunoassay for
the mammalian and nonmammalian assays.[12] In
addition to interest in measurement of steroids in fish plasma as an
aid to aquaculture, there is increasing interest in such measurements
to monitor the effects of endocrine disrupting chemicals on their reproductive
function.[13] A report[14] has
highlighted the need for a simple, rapid, and reliable method for the
measurement of hormones specific to fish and other nonmammalian vertebrates
as the basis for regulatory testing of the effects of potential endocrine
disrupters on wildlife. In this communication, we describe a general
preparation procedure for the enzyme label of carp Vtg, which is applicable
to other species. The method was applied to the determination of Vtg
production by ethynylestradiol, a potent xenoestrogen, in Carp ( Cyprinus carpio )
plasma.
Materials and Methods
Chemicals and equipment
The C. carpio Vtg and anti-Vtg antibodies were kind gifts
from Professor Oliana Carnevali, Dipartimento di Biologia MCA, University
of Camerino, 62032 Camerino, Italy. The purified Vtg is diluted to 10 and
50 mg/ml with Vtg storage buffer [PPBS, 0.1 M, pH 7.4; Potassium chloride,
27 mM; Tween-20, 0.05%; Bovine Serum Albumin, 0.2%; Sodium Azide, 5 mM; Aprotinin, 20 TIU/ml, 1% (Sigma, Deisenhofen, Germany)], divided into various volumes in eppendorf tubes and stored at -20°C. Goat F (ab′)2 fragment of affinity isolated rabbit antigen-specific antibody, conjugated to alkaline phosphatase was obtained from Sigma Immuno Chemicals (catalogue number A-3937). 4-Nitrophenylphosphate (pNPP) obtained from Boehringer Mannheim GmbH (catalogue number 107 905).
Ninety-six well polystyrene high-binding microtiter plates (Costar catalogue
number 3590) were used for the ELISA. Plates were sealed for storage
with Anachem Sealplate sealing film. Plates were processed using a Chelsea
Instruments shaker, an Anthos Model AW-1 plate washer and read with an
Anthos Model HTII Plate Reader. Ninety-six well low-binding microtiter
plates (Costar catalogue number 2501) were used for sample dilution and
preparation of standard curves and AnachemÒ SEALPLATE
plate sealing film, (PK/100) purchased from Anachem (United Kingdom).
Fish exposure
Four groups of six male common carp (initial mean weight: 268.9 ± 52.5 g, mean ± SD) originating from a local fish breeder farm (Mahyar company, Iran) were kept in four separate tanks. They were exposed to 0, 5, 10, and 50 ng/ml ethynylestradiol for 30 days. Blood samples were taken at 0, 10, 20, and 30 days of exposure and plasma kept at -20°C
until Vtg assay.
Plate coating with Vtg
ELISA plates were coated with 50 ng of purified Vtg. About 100
ml of carbonate buffer (0.05 M, pH 9.6) containing 500 ng/ml of purified
standard Vtg added to each well of the plate and incubated for 1 h at room
temperature in a humid container and then overnight at 4°C and washed once with nanopure water. The plate was blocked by the addition of a 200 ml per well of fresh Vtg blocking buffer [potassium phosphate buffer (PPB) 0.1 M, pH 7.4; bovine serum albumin, 2%; sodium azide, 5 mM], washed three times with wash buffer [Potassium phosphate buffered saline (PPBS), 0.02 M, pH 7.4; Tween-20, 0.05%] sealed with plate cover and stored at -20°C.
Standard assay procedure
Serial dilutions of purified standard Vtg (2000 - 3.9 ng/ml) in 100
ml Vtg assay buffer were made in the first ten wells of rows A and B of
the coated plate. The remaining two wells in these rows were reserved for
B0 (0pg) and nonspecific bound (NSB). Ten microliters of the diluted samples
from sample dilution plate (2, 200, and 2000 times) was pipetted into the
remaining six rows in duplicate. Twenty microliters of anti-Vtg antisera
diluted 1/15 000 (carp) in Vtg assay buffer was added to all wells apart
from NSB wells and made up the wells to 150 ml by Vtg assay buffer. The
plates were sealed and incubated 2 h at room temperature in a humid chamber.
Plates were then washed three times with wash buffer and 150 ml of anti-rabbit
IgG alkaline phosphatase conjugate diluted 1/15 000 in Vtg assay buffer
added to each well and incubated overnight at 4°C temperature. Plate
was washed with wash buffer once and then twice with pNPP Substrate buffer
(diethanolamine, 0.5 M, pH 9.8; MgCl, 0.5 mM), 175 ml/well of 1 mg/ml 4-nitrophenylphosphate
in pNPP Substrate buffer added and then incubated in dark at room temperature
until color developed to around 1.5 ODs (normally approximately 45 min
but can be left up to 4 h). The reaction stopped by adding 50 ml of 3 M
NaOH and read at 405 nm. Nanograms per ml were calculated for the samples
from the standard curve using Stingray software (Dazdaq, Ringmer, UK).
Assay validation
A pool of male common carp ( C. carpio ) plasma (500 ml) was
treated with 100°C for 1 h to remove all proteins from the plasma.
One hundred microliter aliquots were pipetted in duplicate into wells on
two rows of a microtiter plate. Pure Vtg was added to one pair of wells
to give a concentration of 2000 ng/well, serial dilutions (2x) performed
and the samples assayed according to standard procedure. The same procedure
was used to assay Vtg in by carp exposed to ethynylestradiol (0, 5, 10,
and 50 ng/ml). SPSS 10 for Windows software (SPSS Inc., 444 N. Michigan
Avenue, Chicago, Illinois 60611, USA) was used for the statistical analysis
of Vtg production in plasma of control fish group (0 ng/ml ethynylestradiol)
with other experimental groups (5, 10, and 50 ng/ml ethynylestradiol).
Results
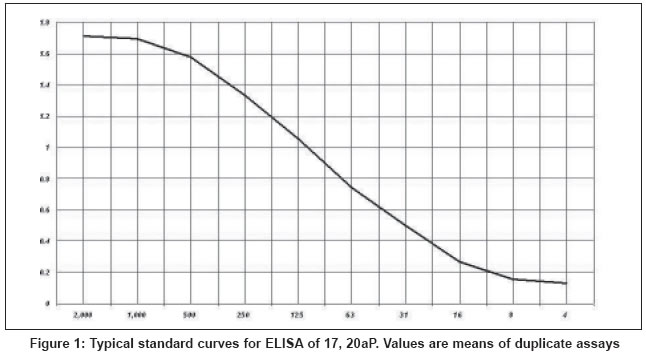
Standard curves and assay sensitivity
Typical standard curves for Vtg showed the working range of the ELISA was 11.25 - 2000 ng/ml [Figure
- 1] [75
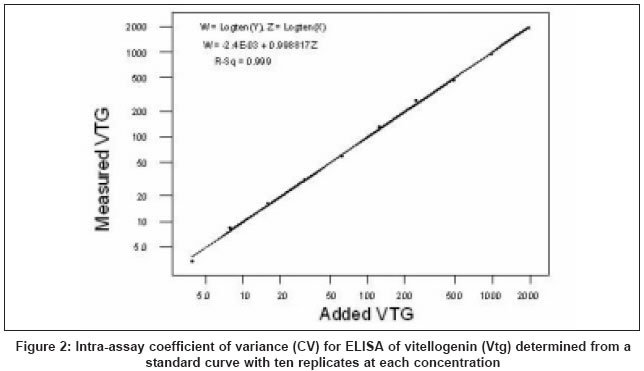
- 25% specific binding/maximum antibody binding (B/B0)] with a 50% B/B0 intra and interassay variation of 3.9% ( n = 10) [Figure
- 2] and
12.5% ( n = 30), respectively. This ELISA is capable of detecting Vtg as low as 6 ng/ml.
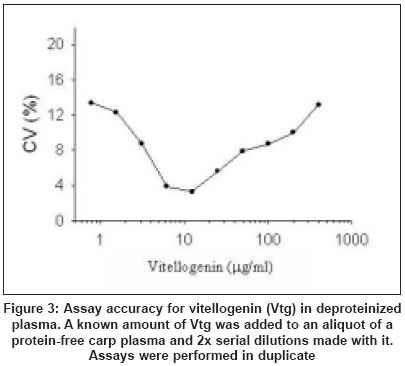
Assay accuracy
There was a strong positive correlation ( r > 0.999) between
the amount of Vtg added to deproteinated plasma and the amount found [Figure
- 3].
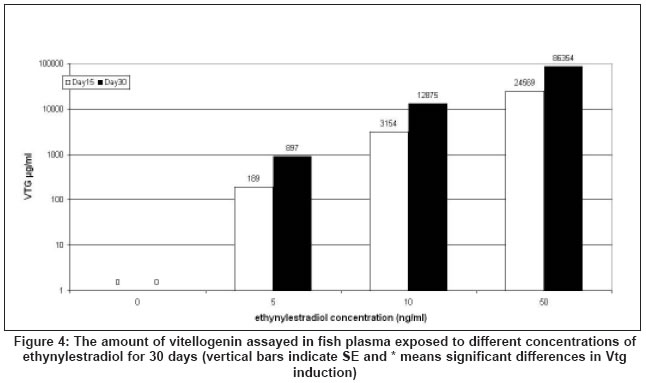
Application to measuring Vtg production by carp exposed to ethynylestradiol
The Vtg was measured in duplicate by ELISA using 10 ml of the diluted
plasma (2x, 200x, and 2000x) from fish exposed to different concentrations
of ethynylestradiol. The amounts of Vtg detected by this method was 0,
402 ± 50.6, 6790 ± 1235, and 49824 ± 10423 mg/ml (mean ± standard
error of mean, for six fish in each group) for fish exposed to 0, 5, 10,
and 50 ng/ml of ethynylestradiol for 30 days respectively [Figure
- 4].
A clear and potent estrogenic activity in inducing Vtg by fish liver was
found with increase in ethynylestradiol concentration exposure and the
amount of Vtg induced by different concentrations of ethynylestradiol between
control (0 ng/ml) and experimental (5, 10, and 50 ng/ml) groups and between
experiment groups were different significantly ( P < 0.05).
Discussion
Examination for the effects of potential EDCs begins only at levels at which there is no apparent stress or discomfort to the animal, so that it is potentially and equally able to reproduce.[15] Reproductive dysfunction at such low concentrations can be caused either by direct action on the gametes, or indirectly by modulation of the endocrine system so that gamete development takes place during an imbalance in the hormonal environment.[16]
In female fish, endocrine modulators can affect hypothalamic gonadotrophin releasing hormone (GnRH) secretion, pituitary gonadotrophin (GtH) release, estrogen biosynthesis and catabolism, estrogen binding to hepatic receptors, hepatic Vtg production, or the feedback by steroids and Vtg.[17] Although hepatic production of Vtg in male fish could be initiated by estrogens hormone, but no estrogenic hormones secrete in male fish and no Vtg production can be expected in normal life.[7] The use of in vitro fish hepatocyte cultures or in vivo fish exposure to estrogenic chemicals is shown to provide a simple and effective way to screen estrogenic activities of EDCs.[18] The Vtg assay in cell culture or fish plasma needs a quick, cheap, reliable, and efficient method to fulfill such a duty[19] and many researchers have already introduced different techniques to measure Vtg.[20],[7] An ELISA method for assaying Vtg has been described here.
Assay validation
Sensitivity: Sensitivity of the assays, which has been taken
as 90% of
the binding with 0pg is just over 10 mg for Vtg. Using identical conditions
for label preparation and assay we also obtained a sensitivity of 2 mg
for Vtg comparable to that obtained by others.[21] As
with radioimmunoassay, the sensitivity is a function of both the specific
activity of the label and the quality of the antiserum, and will therefore
vary with the material available to the user. So in this study, we used
the antiserum that were available to us but have little reason to expect
that any antiserum which gives good results with RIA will not give similar
results to those that we describe for ELISA.
Previous reports of ELISA for Vtg have used different enzyme labels[22] and
in this communication a method described previously[23] and
the results showed it is sufficiently sensitive for the rapid measurement
of Vtg in common fish species such as cyprinids, and an example of such
an application was described.
Accuracy and reproducibility: A major advantage of ELISA is the
ability to minimize errors resulting from multiple pipetting by use of
multichannel pipettes. Inter and intra-assay coefficient of variations
(CV) reported in RIA are frequently 15-20% even when measured at the steepest part of the curve. For the assays reported here we obtain intra-assay CVs around 3.9% in the steep part of the curve, rising to 15-20% at
the flatter end regions [Figure - 2],
while interassay variation (at 50% displacement) was around 12%,
comparable to that reported by Cuisset et al.[23] for
11-ketotestosterone steroid hormone. There was a highly significant correlation
between the Vtg added to and that measured in deproteinized carp plasma [Figure
- 3] and
carp plasma. A further advantage of ELISA is that sample handling can also
be made more rapid and reproducible by the use of low binding microtiter
plates for sample storage and dispensing with multichannel pipettes
Application to the measurement of Vtg production in fish
To demonstrate an application of the ELISA method we have measured
the amount of Vtg produced by carp exposed to ethynylestradiol. The results
confirmed the previous findings that ethynylestradiol is a potent estrogenic
chemical[24] [Figure
- 4] and
a clear, and potent estrogenic activity in inducing Vtg by fish was found
with increase in ethynylestradiol concentration and the time of exposure
(15 vs 30 days) [Figure - 4].
The amounts of Vtg production in fish exposed increased dramatically
so that the logarithmic scales were used to demonstrate it as shown in [Figure
- 4].
Such a potent induction of Vtg in other studies have already been discussed.[25]
The rapid determination of Vtg from the very small volumes of blood allows
the examination of water pollution with EDCs and this methodology will
therefore be of great interest to workers screening the estrogenic of
new chemicals and phytoestrogens.
References
| 1. | Tsutsumi O. Effects of endocrine disruptors on preimplantation embryo development . Nippon Rinsho 2000;58:2464-8. Back to cited text no. 1 [PUBMED] |
| 2. | Damstra T, Barlow S, Bergman A, Kavlock S. Global assessment of the state of the science of endocrine disruptors . 2002;30-60. Back to cited text no. 2 |
| 3. | Lathers CM. Endocrine disruptors: a new scientific role for clinical pharmacologists? Impact on human health, wildlife, and the environment . J Clin Pharmacol 2002;42:7-23. Back to cited text no. 3 [PUBMED] [FULLTEXT] |
| 4. | Nakari T. Estrogenicity of municipal effluents assessed in vivo and in vitro . Environ Toxicol 2004;19:207-15. Back to cited text no. 4 [PUBMED] [FULLTEXT] |
| 5. | Versonnen BJ, Janssen CR. Xenoestrogenic effects of ethinylestradiol in zebrafish (Danio rerio) . Environ Toxicol 2004;19:198-206. Back to cited text no. 5 [PUBMED] [FULLTEXT] |
| 6. | Hornung MW, Jensen KM, Korte JJ, Kahl MD, Durhan EJ, Denny JS, et al . Mechanistic basis for estrogenic effects in fathead minnow (Pimephales promelas) following exposure to the androgen 17alpha-methyltestosterone: conversion of 17alpha-methyltestosterone to 17alpha-methylestradiol . Aquat Toxicol 2004;66:15-23. Back to cited text no. 6 |
| 7. | Aerni HR, Kobler B, Rutishauser BV, Wettstein FE, Fischer R, Giger W, et al . Combined biological and chemical assessment of estrogenic activities in wastewater treatment plant effluents . Anal Bioanal Chem 2004;378:688-96. Back to cited text no. 7 |
| 8. | Sharpe RL, MacLatchy DL, Courtenay SC, Van Der Kraak GJ. Effects of a model androgen (methyl testosterone) and a model anti-androgen (cyproterone acetate) on reproductive endocrine endpoints in a short-term adult mummichog (Fundulus heteroclitus) bioassay . Aquat Toxicol 2004;67:203-15. Back to cited text no. 8 [PUBMED] [FULLTEXT] |
| 9. | Versonnen BJ, Goemans G, Belpaire C, Janssen CR. Vitellogenin content in European eel (Anguilla anguilla) in Flanders, Belgium . Environ Pollut 2004;128:363-71. Back to cited text no. 9 [PUBMED] [FULLTEXT] |
| 10. | Madsen LL, Korsgaard B, Bjerregaard P. Estrogenic effects in flounder Platichthys flesus orally exposed to 4-tert-octylphenol . Aquat Toxicol 2003;64:393-405. Back to cited text no. 10 [PUBMED] [FULLTEXT] |
| 11. | George S, Gubbins M, MacIntosh A, Reynolds W, Sabine V, Scott A, et al . A comparison of pollutant biomarker responses with transcriptional responses in European flounders (Platicthys flesus) subjected to estuarine pollution . Mar Environ Res 2004;58:571-5. Back to cited text no. 11 |
| 12. | Yamada H, Satoh R, Ogoh M, Takaji K, Fujimoto Y, Hakuba T, et al . Circadian changes in serum concentrations of steroids in Japanese char Salvelinus leucomaenis at the stage of final maturation . Zoolog Sci 2002;19:891-8. Back to cited text no. 12 |
| 13. | Wade MG, Foster WG, Younglai EV, McMahon A, Leingartner K, Yagminas A, et al . Effects of subchronic exposure to a complex mixture of persistent contaminants in male rats: systemic, immune, and reproductive effects. Toxicol Sci 2002;67:131-43. Back to cited text no. 13 |
| 14. | Tattersfield L, Matthiessen P, Campbell P, Grandy N, Lange R. SETAC-Europe/OECD/EC Expert Workshop on Endocrine Modulators and Wildlife: Assessment and Testing (EMWAT) . Proceeding of SETAC-Europe/OECD/EC Expert Workshop: Veldhoven, The Netherlands, SETAC-Europe, Amsterdam; 1997. Back to cited text no. 14 |
| 15. | Ackermann GE, Schwaiger J, Negele RD, Fent K. Effects of long-term nonylphenol exposure on gonadal development and biomarkers of estrogenicity in juvenile rainbow trout Oncorhynchus mykiss . Aquat Toxicol 2002;60:203-21. Back to cited text no. 15 [PUBMED] [FULLTEXT] |
| 16. | Okai Y, Sato EF, Higashi-Okai K, Inoue M. Enhancing Effect of the Endocrine Disruptor para-Nonylphenol on the Generation of Reactive Oxygen Species in Human Blood Neutrophils . Environ Health Perspect 2004;112:553-6. Back to cited text no. 16 [PUBMED] [FULLTEXT] |
| 17. | Berg AH, Westerlund L, Olsson PE. Regulation of Arctic char (Salvelinus alpinus) egg shell proteins and vitellogenin during reproduction and in response to 17beta-estradiol and cortisol . Gen Comp Endocrinol 2004;135:276-85. Back to cited text no. 17 [PUBMED] [FULLTEXT] |
| 18. | Tyler CR, Pottinger TG, Santos E, Sumpter JP, Price SA, Brooks S, et al . Mechanisms controlling egg size and number in the rainbow trout, Oncorhynchus mykiss . Biol Reprod 1996;54:8-15. Back to cited text no. 18 |
| 19. | Fujita T, Fukada H, Shimizu M, Hiramatsu N, Hara A. Quantification of serum levels of precursors to vitelline envelope proteins (choriogenins) and vitellogenin in estrogen treated masu salmon, Oncorhynchus masou . Gen Comp Endocrinol 2004;136:49-57. Back to cited text no. 19 [PUBMED] [FULLTEXT] |
| 20. | Anderson MJ, Cacela D, Beltman D, Teh SJ, Okihiro MS, Hinton DE, et al . Biochemical and toxicopathic biomarkers assessed in smallmouth bass recovered from a polychlorinated biphenyl-contaminated river . Biomarkers 2003;8:371-93. Back to cited text no. 20 |
| 21. | Volz DC, Chandler GT. An enzyme-linked immunosorbent assay for lipovitellin quantification in copepods: a screening tool for endocrine toxicity . Environ Toxicol Chem 2004;23:298-305. Back to cited text no. 21 [PUBMED] |
| 22. | Martin-Diaz ML, Bambe, S, Casado-Martine C, Sale D, DelValls TA. Toxicokinetics of heavy metals from a mining spill using Carcinus maenas . Mar Environ Res 2004;58:833-7. Back to cited text no. 22 |
| 23. | Cuisset B, Pradelles P, Kime DE, Kuehn ER, Babin P, Davail S, et al . Enzyme immunoassay for 11-ketotestosterone using acetylcholinesterase as label: Application to the measurement of 11-ketotestosterone in plasma of Siberian sturgeon . Comparative Biochemistry and Physiology C Pharmacology. Toxicol Endocrinol 1994;108:229-41. Back to cited text no. 23 |
| 24. | Craft JA, Brown M, Dempsey K, Francey J, Kirby MF, Scott AP, et al . Kinetics of vitellogenin protein and mRNA induction and depuration in fish following laboratory and environmental exposure to oestrogens . Mar Environ Res 2004;58:419-23. Back to cited text no. 24 |
| 25. | Versonnen BJ, Arijs K, Verslycke T, Lema W, Janssen CR. In vitro and in vivo estrogenicity and toxicity of o-, m-, and p-dichlorobenzene . Environ Toxicol Chem 2003;22:329-35. Back to cited text no. 25 [PUBMED] |
Copyright 2005 - Indian Journal of Occupational and Environmental Medicine
The following images related to this document are available:
Photo images
[oe05016f2.jpg]
[oe05016f1.jpg]
[oe05016f4.jpg]
[oe05016f3.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}