 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
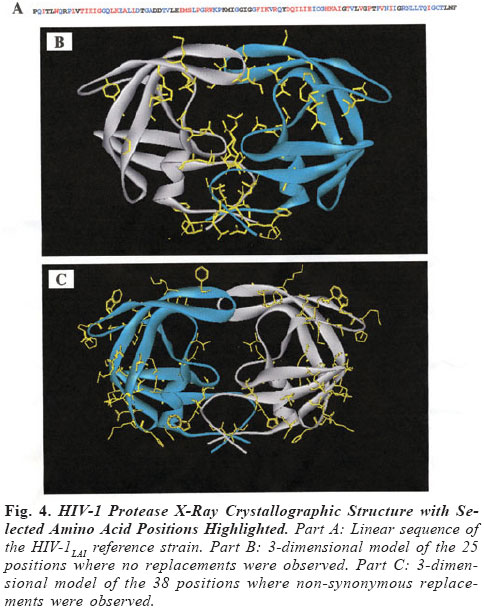
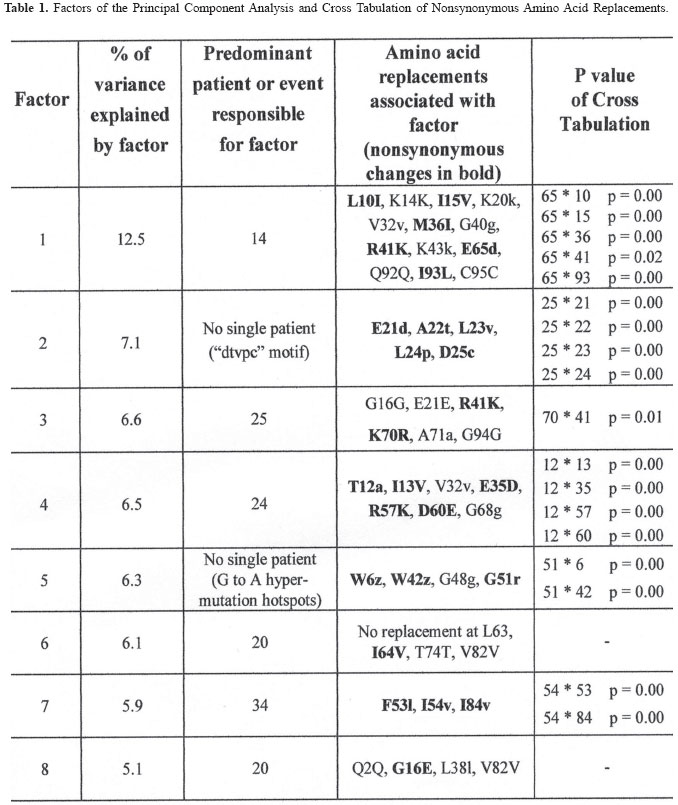
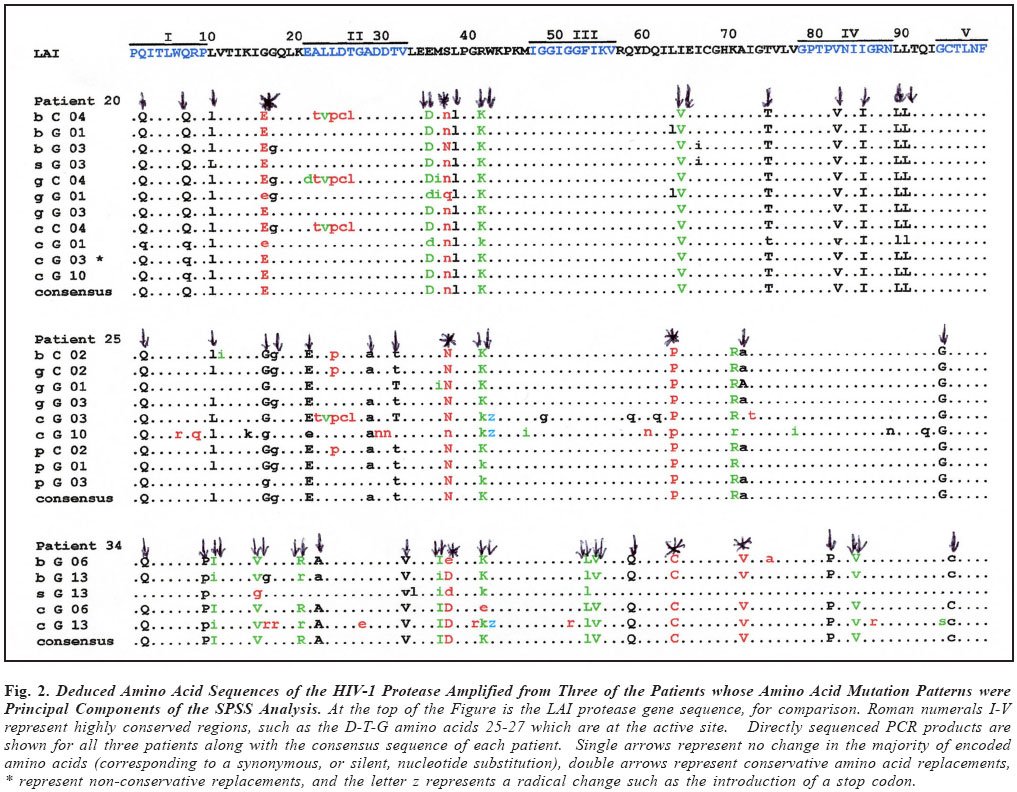
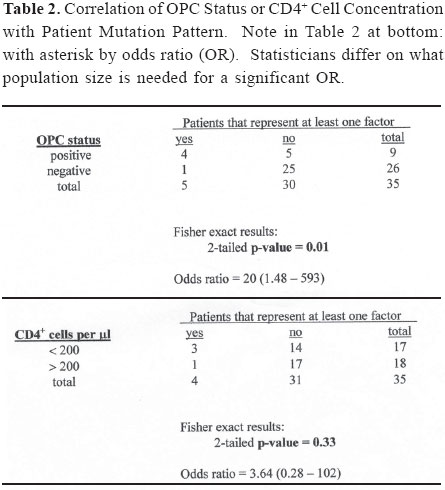
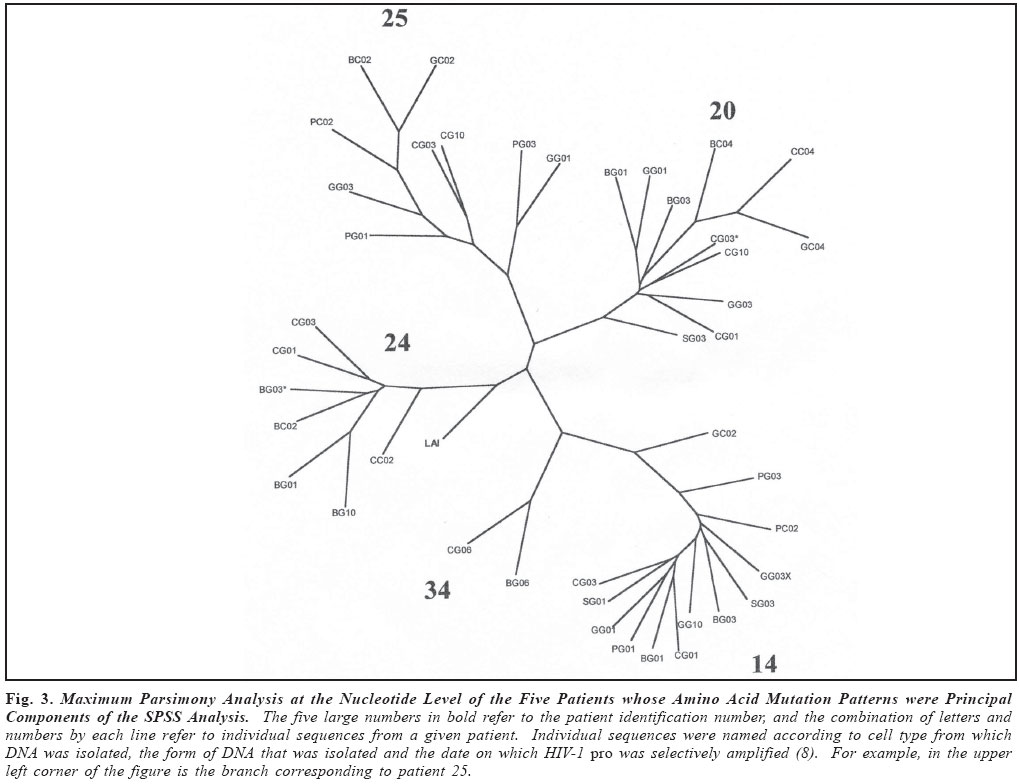
Brazilian Journal of Oral Sciences, Vol. 3, No. 9, April/June 2004, pp. 446-453 Specific protease mutation patterns are associated with oropharyngeal candidiasis in a New Orleans patient cohort of HIV-infected individuals Peter J. Hickman1 Robertino M. Mera2 Janet E. Leigh1,3 Paul L. Fidel Jr.1 Allen R. Mock1 William R. Gallaher1 Ronald B. Luftig1 1Department of Microbiology, Immunology and Parasitology 2Department of Pathology 3Department of General Dentistry Louisiana State University Health Sciences Center Correspondence to: Ronald B. Luftig Louisiana State University Health Sciences Center 1901 Perdido Street New Orleans, LA 70112-1393 Phone: (504)568-4063 Fax: (504)568-2918 e-mail: rlufti@lsuhsc.edu Received for publication: January 31, 2004 Code Number: os04013 Abstract HIV-1 protease gene sequences were obtained from peripheral blood, saliva or oral tissues of 35 HIV+ patients using nested amplification and manual sequencing of PCR products. Of the 35 HIV+ patients 9 had clinical oropharyngeal candidiasis (OPC) while 26 did not, and only 4 patients were on protease inhibitor (PI) therapy. These patients were collected prior to major use of HAART therapy in New Orleans, Louisiana. Analysis of 172 amino acid sequences revealed unique patterns of mutation that were in most cases independent of the type of cell from which DNA was isolated and were, instead, primarily dependent on the individual patient. Principal component analysis indicated that approximately 50% of the variance of the amino acid replacements could be explained by patterns of change seen in only five patients. Significantly, 4 of these 5 patients were OPC+ indicating that patients with OPC are more likely to express a principal mutation pattern than patients without OPC (p = 0.002, Chi square). Dendrograms revealed that these five patients clustered separately from each other and from HIV-1LAI suggesting that principal mutation patterns as well as OPC are independent of viral evolution. In conclusion, prior to widespread use of PI therapy to combat HIV-1, patients with OPC exhibited unique patterns of amino acid replacements within the HIV-1 protease. Key Words: HIV-1, mutation, proteases, candidiasis Introduction One of the key areas of concern for infected patients is the maintenance of a persistent HIV-1 infection in the presence of an active immune response. This is in part due to reservoirs of HIV-1 infection1 wherein the virus is protected from not only host defenses but also from highly active antiretroviral therapy (HAART) (see review by Ogden and Flexner2, 2001). In these sites, the virus or its proviral genome can remain in a latent or persistent state where it can undergo mutational changes in a prolonged infection. Examples of such sites are the oral cavity despite the presence of secretory leukocyte protease inhibitor (SLPI)3, inducible lymphoid sites during HAART4 and quiescent T cells that can be activated with prostatin5. The presence of virus in such reservoirs is evidenced by the rapid rate of viral rebound once HAART or other antiviral therapy is removed6. Despite the relative deficit of virus in the oral cavity, oral lesions such as OPC (oropharyngeal candidiasis), hairy leukoplakia and periodontal diseases (necrotizing gingivitis and periodontitis) as well as human papilloma viral warts have been associated with HIV-1 infection7. The oral carriage rate of Candida sp. is high (60-70%), yet few if any healthy individuals develop oropharyngeal candidiasis. In contrast, the majority of HIV-1 infected individuals, prior to the widespread use of HAART, developed OPC7. This study used programs from the Statistical Package for Social Sciences (SPSS) and the Phylogeny Inference Package (PHYLIP) to examine the relationship of protease gene mutation patterns in a cohort of 35 HIV-1 infected individuals from New Orleans. Based on interviews of patients and examination of their medical records, most of the patients (89%) in this prospective study were not taking protease inhibitors at the time tissue was obtained. Although in 89% of the patients there is no evidence of medically accepted anti-HIV-1 therapy, we cannot rule out alternative medicines or other non-standard therapy. Surprisingly, in this study a high correlation was found with specific mutation patterns in a subset of patients and the presence of OPC. Of course, one cannot eliminate the possibility of spontaneous mutations; however, one would think this unlikely to the degree seen for such a highly conserved gene (see Figure 4 legend). Material and Methods Isolation of template DNA, amplification, and purification and sequencing of PCR products. New Orleans patients were enrolled in this study over a 21 month period only after informed consent, and procedures were performed in strict adherence to the guidelines of the Institutional Review Board of the Louisiana State University Health Sciences Center. The 35 patient cohort was obtained relatively early in the HIV disease epidemic in New Orleans (between 9/30/96 and 6/30/ 98), and only 4 patients (=11.4% of the cohort) were being treated with protease inhibitors as part of HAART. Details on the make-up of this pilot population (ethnicity, sex, age) were taken into account during our statistical analysis and have stood out as confounders. Genomic DNA or cDNA was isolated from peripheral blood mononuclear cells (PBMC or b), unstimulated saliva (s) or oral biopsy of gingival cuff (g), buccal mucosa (c), tongue (t) or palate (p) tissues as described previously8. DNA samples were amplified using nested primers specific for the HIV-1 protease as described previously8. PCR products of the predicted size were gel purified from extraneous bands and excess primers using the QIAquick Gel Extraction Kit (Qiagen, Valencia, CA, USA) according to manufacturer's instructions, and PCR products were directly sequenced using the Thermo Sequenase Radiolabeled Terminator Cycle Sequencing Kit (USB Corporation, Cleveland, OH, USA) as described previously8. Statistical analysis of cohort data. We initially chose to examine the 172 protease sequences by principal component analysis using the Statistical Package for Social Sciences (SPSS) to see whether particular mutation patterns were correlated with source of DNA (genomic DNA versus cDNA), cell type from which the DNA originated (oral compartment versus humoral/systemic) or individual patients. Principal component analysis is a multi-dimensional method that produces a single unique solution from a correlation matrix. Initially, the analysis produces a crude table of variances (not unlike that seen in Table 1) and is followed by a Varimax Rotation of data and interpretation of the solution. Each variable will now have a certain loading value (between -1.0 and 1.0) that describes the correlation between the individual variables and the overall set of factors. The "most significant" scores are listed by factor and the Varimax Rotation results in a fairly even distribution of loading scores9. It is true that the use of SPSS is normally limited by such a small sample size; however, because the experienced statistician on the study (Dr. Robertino Mera) was so impressed that the sum of variances of only five factors approached 50%, we decided to proceed with the analysis and report our results. All nucleotide sequences were translated into amino acid sequences using the universal genetic code and conventional single-letter abbreviations for amino acids. A capital letter indicates that the only species present in the given PCR product is different from the HIV-1LAI reference strain at that position, whereas a small letter indicates a mixture of mutant protease and HIV-1LAI-like protease at the DNA level. When microheterogeneity was observed two or three times within a single codon, we assumed that all mutations were occurring simultaneously. Amino acid substitutions were identified as structurally/functionally conservative, nonconservative or radical replacements according to the system of Gallaher et al.10. In order to increase the signal to noise ratio in the sequence data, amino acid positions were selected for principal component analysis by a three step process as follows: 1) accept only positions where 10% or more of the samples had a replacement; 2) add amino acid positions which by visual inspection appear to be sites of replacement for a particular patient even though they may be rarely-occurring sites of replacement in the overall sample set; 3) perform a principal component analysis and drop all positions that are non-loading (that is, not a component of the first ten factors). The use of PHYLIP programs to produce dendrograms is summarized as a flow diagram in Figure 1. For parsimony analysis, files representing nucleotide sequences were manually adjusted to Pearson's (FastA)11 sequential format using Microsoft Word 97, version SR-2 and Word Pads to produce the text file infilen.txt. DNA and protein sequences were aligned via the CLUSTALW version 1.7 program using the IUB DNA weight matrix or the BLOSUM30 protein weight matrix12. This produced the files "infilen.phy" or "infilep.phy". Nucleotide sequences were translated manually using the universal genetic code and aligned by visual inspection. Protein sequences and alignments were verified using the dnaSP program version 3.0 program to translate "infilen.phy" followed by alignment using the Advanced BLAST version 2.0.8 program13. ResultsIn a more recent 2002 study involving a larger cohort of 80 patients 71% of whom were undergoing antiretroviral, including HAART therapy, we noted that the protease gene encoded amino acid sequences that by visual inspection were unique for each patient8. Furthermore, a "dtvpc" motif at the active site occurred in 6.4% of the protease gene sequences, and a stop codon ("z") was observed in 17.7% of the protease gene sequences. We predict that both of these types of mutation, if expressed, would inactivate the protease and potentially lead to defective viral particles. These unique features are illustrated by the HIV-1 protease sequences of three patients shown in Figure 2. Note that by visual inspection of any given patient there is a high degree of intrapatient similarity in the sequences derived from different cell types, yet the consensus sequences of the three patients in Figure 2 clearly show interpatient differences. SPSS factor analyses revealed unique patterns of amino acid mutations that were mostly dependent on individual patients. The pattern and number of mutations were independent of source of DNA (genomic DNA versus cDNA) and in approximately 90% of the patients were independent of the cell type from which the DNA originated (PBMC, saliva or oral tissues). Cross tabulations of amino acid replacements were performed on 172 sequences obtained from the 35 patient cohort, and the results are shown in Table 1. Six of the eight factors can be attributed to mutation patterns within a small subset of patients. Specifically, five patients have been identified whose mutation patterns explain six of the factors and account for 42.7% of the variance in the entire 35 patient data set. Factor 2 (accounting for 7.1% of the variance) is the frequently occurring "dtvpc" motif near the active site of the protease, and factor 5 (accounting for 6.3% of the variance) is probably the result of guanosine to adenosine mutation hot spots occurring at the only two tryptophan residues of the HIV-1 protease (leading to stop codons) and also occurring in the conserved glycine-rich region corresponding to the flap of the homodimer HIV-1 protease which allows (or precludes) substrate binding. Cross tabulations of nonsynonymous replacements within each factor were calculated by Chi-square analysis. The p values for these tests are shown in the last column of Table 1, and the low p values (< 0.05) indicate that the nonsynonymous amino acid mutations that loaded highest in the factor analysis were significant meaning that they occurred with a frequency higher than expected by chance. We observed that, despite a predominantly OPC-cohort, among the five patients whose amino acid mutation patterns were principal components of the SPSS analysis only patient 24 was OPC-. Fisher statistical analysis was performed to determine the relationship between OPC status or CD4+ cell concentration and the five patients who demonstrate principal mutation patterns (see Table 2). Our data suggest that patients with OPC are significantly more likely to express a principal mutation pattern than patients without OPC (p = 0.01); in contrast, patients with fewer than 200 CD4+ cells/µl are no more likely to express a principal mutation pattern than patients with more than 200 CD4+ cells/µl (p = 0.33). These relationships were verified by Chi square analysis where p values were 0.002 or 0.06, respectively, for OPC+ status or CD4+ cell concentration. Nucleotide sequences were analyzed by common phylogenetic and polymorphic methodologies to examine by another technique the results of our SPSS statistical analysis. A key difference in these approaches is that principal component analysis provides structural relationships among the patients while the PHYLIP programs are designed to reveal epidemiological relatedness among the patients, 80% of whom provided samples within the first five months of the study. In order to provide such an independent statistical approach, an examination of the 172 nucleotide sequences were undertaken by PHYLIP analysis14. This involved performing highly stringent multiple sequence alignments at the nucleotide level and included the HIV-1LAI pattern (equivalent to the consensus Los Alamos HIV-1 Group B sequence except for an A to G substitution at position 110 of the pro gene) as the outgroup15. Figure 3 shows the maximum parsimony analysis at the nucleotide level of the five patients that represent the first eight factors shown in Table 1. There is clearly intrapatient clustering of sequences in the phylogenetic tree, a lack of interpatient similarity and a lack of sample site-specific clustering. Finally, based on the crystal structure of the HIV-1 protease homodimer, we were able to model the protein highlighting amino acid positions (Figure 4A) that demonstrated nonsynonymous replacements or demonstrated no replacements. As can be seen in Figure 4B, amino acid positions that never changed tended to be at the active site and at other points where the two subunits interact. Positions that could tolerate nonsynonymous replacements, in contrast, tended to be on the surface of the protein exposed to the aqueous environment (see Figure 4C). Discussion As noted earlier, the multivariate statistical approach of principal component analysis is designed to analyze the protease substitution patterns at the amino acid level and condense data from the original variables into a small set of factors9. In this study, as seen in Table 1, principal component analysis revealed that 5 patients explained about 43% of the variance and two additional factors showed significance as well, one corresponding to a "dtvpc" motif and another corresponding to G to A hypermutations which have been demonstrated to occur at a high rate in HIV-1 genes16. As we learn more about HIV-1 protease inhibitors, we are finding that PI resistance2, co-infections with tuberculosis1, interactions with health foods6 and integration into quiescent T-cells4 are all mechanisms that allow persistence of HIV-1 and eventually movement of proviral DNA into reservoirs where virus can be activated at late stages of AIDS. In addition to learning more about the basic biology of the HIV-1 protease, this study provides an in-depth analysis of the changes in HIV-1 protease patterns that may be happening in a remote location of the body, the oral cavity, where virus can mutate while persisting associated with cells. Here external factors such as SLPI, the SLPI receptor, uncharacterized entities in saliva as well as Candida-specific secreted aspartyl proteases (SAPs)17 may exert selection on the HIV-1 protease. HIV-1 sequences obtained from oral tissues were distinguishable from those obtained from PBMC in approximately 10% of the patients which is consistent with a small but real variation in pro for the oral cavity8. Conversely, HIV-1 sequences obtained from oral tissues were identical to those obtained from PBMC in approximately 90% of the patients which may reflect contamination of oral tissues with blood or may indicate that the predominant oral virus has moved into the PBMC. Thus, when examining HIV-1 gene sequence variation, oral sites should be considered especially in OPC+ patients. Cytokine expression is different in the oral cavity, and candidiasis may also exert a selective pressure in the presence of protease inhibitors18 potentially leading to increased production of variant proteins or defective particles that might be more lethal at advanced stages of disease19. There is also in vitro evidence using a murine model that C. albicans variants with elevated levels of SAPs are more pathogenic than isolates that produce low levels of SAPs20. We do not know what selective pressure SAPs might have on the HIV-1 protease. Since some antiretroviral PIs are known to crossreact with some of the SAPs produced by C. albicans18, it is possible, for example, that high levels of SAPs might compete with HIV1 protease for PI binding and effectively protect the HIV-1 protease from developing PI resistant mutations. On the other hand, perhaps high levels of SAPs compete for viral precursor substrates and interfere with the maturation of viral particles unless particular mutations in the HIV-1 protease emerge. Experiments will be performed where recombinant SAPs are added to HIV-1LAI-infected cells to see if specific mutations arise in the HIV-1 protease. It is unclear whether OPC causes a higher mutation rate and unique mutations to arise in the HIV-1 protease or if specific mutations in the HIV-1 protease affects the immune system in such a way that the patient is more susceptible to OPC infection. Regardless of cause and effect, there is clearly a relationship between OPC and unique patterns of mutation in the HIV-1 protease among patients where there is little selective pressure on the HIV-1 protease due to PI therapy. We are currently investigating the possibility that other oral infections such as oral hairy leukoplakia (OHL) or human papillomavirus (HPV) might interact with HIV-1 in a manner similar to OPC leading to high numbers or unique patterns of mutation in the HIV-1 protease. Finally, Figure 4 shows that the mutations selected in HIV+ patients who have "not" been extensively treated with HAART occur predominantly in regions outside of the highly structurally conserved active site and flap. One cannot rule out that the cluster effect seen for OPC represents transmission from the same partner or intravenous transmission at the same period. We also expect that as patients are increasingly treated with HAART we should see mutations at characteristic sites of PI resistance. AcknowledgmentsThis work was supported in part by NIH grant DE-12178 awarded to Dr. Paul L. Fidel Jr. and institutional funds. We wish to thank Stephen Strickland, Ryan Jackson and Jennifer Abadie for their technical assistance as summer students in the lab of Dr. Ronald B. Luftig. Especially, this work could not have been performed without the extensive mentorship provided to Allen Mock by Dr. William Gallaher. References
Copyright 2004 - Piracicaba Dental School - UNICAMP São Paulo - Brazil |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}