 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Brazilian Journal of Oral Sciences, Vol. 4, No. 12, Jan./Mar. 2005, pp. 651-658
Dental cementum reviewed: development, structure, composition,regeneration and potential functions Patricia Furtado Gonçalves 1 Enilson Antonio Sallum 1 Antonio Wilson Sallum 1 Márcio Zaffalon Casati 1 Sérgio de Toledo 1 Francisco Humberto Nociti Junior 1 1 Dept. of Prosthodontics and Periodontics,
Division of Periodontics, School of Dentistry at Piracicaba - UNICAMP,
Piracicaba, São Paulo, Brazil. Received for publication: October 01, 2004 Code Number: os05003 Abstract This article reviews developmental and structural characteristics of cementum, a unique avascular mineralized tissue covering the root surface that forms the interface between root dentin and periodontal ligament. Besides describing the types of cementum and cementogenesis, attention is given to recent advances in scientific understanding of the molecular and cellular aspects of the formation and regeneration of cementum. The understanding of the mechanisms involved in the dynamic of this tissue should allow for the development of new treatment strategies concerning the approach of the root surface affected by periodontal disease and periodontal regeneration techniques. Key Words: dental cementum, review Introduction
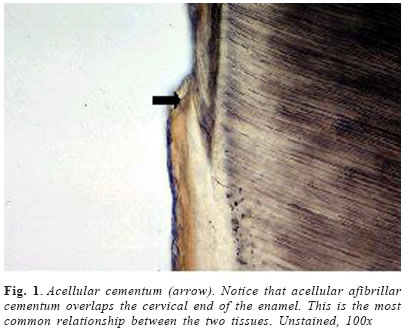
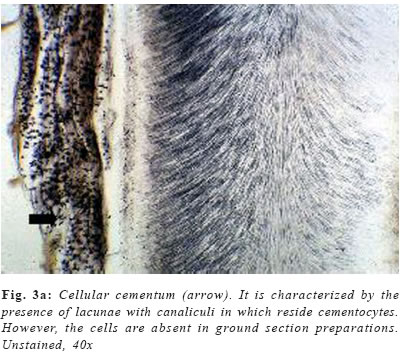
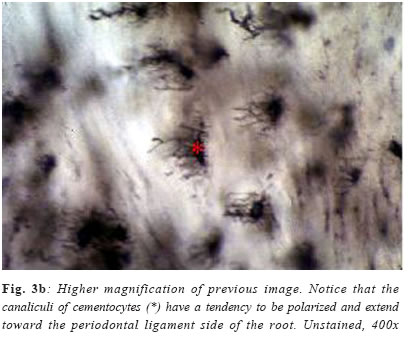
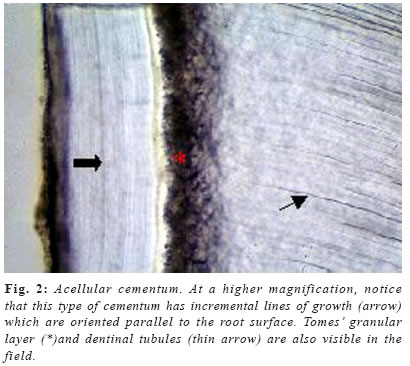
Cementum is an avascular mineralized tissue covering the entire root surface. Due to its intermediary position, forming the interface between root dentin and periodontal ligament, cementum is a component of the tooth itself, but belongs functionally to the dental attachment apparatus, that is, the periodontium. One of the main functions of cementum is to anchor the principal collagen fibers of the periodontal ligament to the root surface, but it also has important adaptative and reparative functions, playing a crucial role to maintain occlusal relationship and to protect the integrity of the root surface. Until recently, knowledge of the development and the general structure of the periodontium was limited to morphological information, although, during the last decade, rapid progress has been made in understanding the molecular and cellular biology of periodontal tissue development and regeneration, including cementum. Dental cementum is unique in various aspects: it is avascular and not innervated, does not undergo continuous remodelling like bone, but continues to grow in thickness throughout life1. In contrast with these specific histological characteristics, it appears not to be specific at the cellular and molecular level2. Unlike dentine and enamel, where there are clear differences in the proteins present in these tissues and the factors regulating their functions when compared with bone, cementum has not demonstrated to express specific proteins, appearing to contain factors in common with bone and to be developmentally controlled by similar factors2-3. There is accumulating histological evidence that cementum is critical for appropriate maturation of the periodontium, both during development and as well as that associated with regeneration of periodontal tissues3. Recent studies have contributed to an understanding of the possible involvement of some of the molecular factors in cementum regeneration4, but cementogenesis, on a cell biological basis, continues to be poorly understood. The aim of this chapter is to give a comprehensive review about some important aspects of this unique tissue, including its varieties, development, structure, composition, regeneration and potential functions. Types of cementum Traditionally, cementum has been classified as cellular and acellular, depending on the presence or absence of cementocytes in its structure. Another classification includes intrinsic or extrinsic fiber cementum, depending on the presence of collagen fibers formed by cementoblasts or by fibroblasts, respectively5-6. Mainly three cementum types differing in these aspects are distinguished in humans. Acellular afibrillar cementum covers minor areas of the enamel, particularly at and along the cementoenamel junction (Figure 1). The areas and location of acellular afibrillar cementum vary from tooth to tooth and along the cementoenamel junction of the same tooth6-9. Its major structural organic components are glycosaminoglycans6, and its functional significance is unknown. The lack of collagen fibrils indicates that this cementum variety has no function in tooth attachment. Cellular intrinsic fiber cementum contains cementocytes embedded in a collagenous matrix of intrinsic collagen fibers (Figures 3a , b and 4). These collagen fibers are oriented mostly parallel to the root surface and course in a circular fashion around the root6. A fast matrix deposition by the cementoblasts, which occurs in the space between deviating epithelial cells of Hertwig´s root sheath and the dentinal surface appears to be the reason for the incorporation of some of the cementoblasts10-11. Cellular intrinsic fiber cementum is found in the furcation, on the apical root portion, in old resorption lacunae and in root fracture sites, playing an important role as an adaptative tissue that brings and maintains the tooth in its proper position and also participating in the repair process, although it has no immediate function in tooth attachment1,6. Only cellular intrinsic fiber cementum can repair a resorptive defect of the root in a reasonable time, due to its capacity to grow much faster than any other type of cementum10. Acellular extrinsic fiber cementum is mainly found on cervical and middle root portions, covering 40% to 70% of the root surface. On anterior teeth, it may also cover part of the apical root portion, since its apical extension increases from posterior to anterior teeth. It serves the exclusive function of anchoring the root to periodontal ligament. The acellular extrinsic fiber cementum matrix consists of a dense fringe of short collagenous fibers that are implanted into the dentinal matrix (glycosaminoglycans) and are oriented about perpendicularly to the root surface6. When they become elongated and eventually continuous with the principal periodontal ligament fibers they are called Sharpey´s fibers. Although acellular extrinsic fiber cementum is a quite constantly growing tissue, appositional lines (Figure 2) represent the periodical deposition of cementum layers in frequent association with an abrupt change in the direction of Sharpey´s fibers12. Moreover, as can be deducted from faster growth rates on distal (4.3mm/year) than on mesial (1.4mm/year) root surfaces13, acellular extrinsic fiber cementum has the potential to adapt to functional alterations such as mesial tooth drift. In humans, the extrinsic fibers traverse and intermingles the intrinsic fiber cementum variety either sporadically or densely arrayed in parallel; this mixed cementum is referred to as cellular mixed stratified cementum6. Origin and development of cementum Based on the observation that populations of cementoblasts are phenotipically distinct, some authors defended the possibility that acellular and cellular cementum have different developmental origin14. Some studies15-16 suggested that cementoprogenitor cells arise from the dental follicle proper, which is of ectomesenchymal origin (that is, a derivative of the cranial neural crest). However, as pointed out by Thomas & Kollar17, labeled cementoblasts could also be of epithelial origin, since cells of the enamel organ, which give rise to HERS, also incorporate 3Hthimidine prior to transplantation in mice. More recent ultrastructural and immunohistochemical studies support, indeed, the hypothesis that cementoblasts originate from epithelial cells of HERS when they undergo an epithelialmesenchymal transformation18-20. There is increasing evidence that Hertwig’s epithelial root sheath is actively involved in the formation of both acellular and cellular cementum. Diekwisch21 (2001), published an extensive literature review on early cementogenesis and performed a detailed morphological and molecular analysis to illustrate and verify key issues in the current debate about epithelial and mesenchymal contributions to root cementum. The author demonstrated that prior to cementogenesis, Hertwig’s epithelial root sheath disintegrates and dental follicle cells penetrate the epithelial layer to invade the root surface. Other studies confirmed that HERS became disrupted or disintegrated prior to cementum deposition, and visualized how mesenchymal cells from the dental follicle penetrated the HERS bilayer and deposited initial cementum, while immediately adjacent epithelial cells were separated from the root surface by a basal lamina and did not secrete any cementum22-23. Human specimen from the Gottlieb collection indicated that HERS was removed from the root surface prior to cementum deposition. In situ hybridization and immunolocalization data revealed that both amelogenin mRNAs and enamel proteins were restricted to the crown enamel and were absent from the root surface and from the cervical-most ameloblasts adjacent to the root margin24-26. On Western blots, cementum protein extracts did not crossreact with amelogenin antibodies25. These studies in conjunction with a literature review together confirmed the theory of cementum as a dental follicle derived connective tissue that forms subsequent to HERS disintegration. Briefly, cementogenesis begins with the deposition of a matrix on the dentin surface by Hertwig’s epithelial root sheath, disruption of the HERS, migration and organization by ectomesenchymal cells from the dental papilla, and their subsequent differentiation into cementoblasts25,27. The matrix they produce surrounds cells producing cementum, the cementoblasts, and the cementoblasts are generated by differentiation of progenitor cells, which in turn are believed to arise from the dental follicle. The fate of HERS following the onset of cementogenesis is also controversial. Traditional thinking proposed that HERS disintegrated into small clusters and/or strands of epithelial cells that survived indefinitely in the periodontal ligament. More recent studies have suggested that epithelial cells of the Hertwig’s root sheath undergo epithelial /mesenchymal transformation into fibroblasts and cementoblasts, that deposit acellular and cellular cementum, respectively1,28. The possibility that some epithelial cells of the root sheath undergo epithelial/mesenchymal transformation and subsequently secrete cementum matrix must be investigated further. There is evidence that cells of the inner layer of the root sheath become incorporated in cellular cementum or trapped between cementum and dentin during formation of the apical part of the root29. The only incontrovertible fact is that many cells of the HERS retain an epithelial phenotype, and survive in periodontal ligament as the epithelial rests of Malassez1,6. Composition
Since cementum is not a uniform, mineralized connective tissue, differences in the proportional composition of the chemical constituents exist between the cementum varieties. Thus, the percentages of its chemical components may vary from sample to sample, particularly in different species. Biochemical studies have shown that cementum has a similar composition to bone. To about equal parts per volume, cementum is composed of water, organic matrix and mineral. About 50% of the dry mass is inorganic, and consists of hydroxyapatite crystals. The remaining organic matrix contains largely collagens, glycoproteins and proteoglycans1. Mineral composition
Cementum is generally less mineralized than root dentin from the same teeth30-31, although not without exception32. Acellular extrinsic fiber cementum appears more highly mineralized than cellular intrinsic fiber cementum and cellular mixed stratified cementum33, what can in part be explained by the presence of uncalcified spaces, such as lacunae and by the uncalcified core of Sharpey’s fibers. Chemical analysis and physicochemical studies indicate that the mineral component is the same as in other calcified tissues; that is, hydroxyapatite (Ca10(PO4)6(OH)2), with small amounts of amorphous calcium phosphates present34. Due to its lower crystallinity of the mineral component (compared to other hard tissues), cementum has a greater capacity for adsorption of fluoride and other elements over time but also more readily decalcifies in the presence of acidic conditions1. As in other hard tissues, the hydroxyapatite of cementum is not pure, but contains other elements (ions) incorporated into mineral phase during mineralization, depending on their concentration in the fluid environment. Over time, the concentration may change by additional uptake or substitution by other ions. Thus, cementum contains 0.50.9% magnesium35-36, about half that in dentin, and it is lower at the surface than in deeper layers of cementum. The significance of the low Mg+2 content in cementum remains obscure, but agrees with the notion that the composition of cementum is more similar to bone tissue than to dentin. In contrast, the distribution of fluoride shows the opposite trend. Cementum have a high fluoride content compared to other mineralized tissues (up to 0.9% ash weight), and this concentration shows a general increase with age and vary with fluoride supply to the individual, as it is in bone, dentin and enamel as well37. Cementum also contains 0.1-0.3% sulfur as a constituent of the organic matrix38, and a number of trace elements may be present in concentrations detectable by electron microprobe analysis, in particular Cu+2, Zn+2 and Na+2 36; however, their distribution and significance do not seem to have been studied in detail. Organic composition
The organic matrix of cementum is composed primarily of collagens. Type I collagen plays structural as well as morphogenic roles and provides scaffolding for mineral crystals; it is the major component, accounting for 90% of all collagens. The type III collagen, which coats type I collagen fibrils, accounts for only 5%3. Cementum contains two major non-collagenous proteins, bone sialoprotein (BSP) and osteopontin (OPN). Both are phosphorilated and sulfated glycoproteins. These proteins, which are prominently expressed in acellular extrinsic fiber cementum and acellular afibrillar cementum, bind tightly to the collagenous matrices and hydroxiapatite, and they possess cell attachment properties through the Arg-Gly-Asp (RGD) sequence, that binds to integrins39-40. Both proteins are expressed during early tooth root development by cells along the root surface. Root surface cells express the BSP, and it is also present in mature teeth. In contrast, OPN is present within the periodontal ligament region of the mature teeth. These two proteins are believed to play a major role in the differentiation of the cementoblasts progenitor cells to cementoblasts3. The BSP is believed to have adhesion function to root surface cells and to participate in initiating mineralization. It is chemotatic to pre-cementoblasts and promotes their adhesion and differentiation41. Many cells express the OPN during periods of cementogenic activity. It regulates cell migration, differentiation, and survival through the interaction with integrins, and also participates in inflammation by regulating monocyte-macrophage activation, phagocytosis, and nitric oxide production. In teeth and cementum, it may regulate biomineralization by at least two mechanisms: regulating bone cell differentiation and matrix mineralization42. Fibronectin43, which is believed to bind cells to the extracellular matrix, and tenascin are present in HERS during odontoblast differentiation, and later at the attachment site of periodontal ligament to the root surface. Osteonectin, osteocalcin and laminin are other matrix components found in cementum. Osteonectin is expressed by cementoblasts producing cellular extrinsic fiber cementum and cellular intrinsic fiber cementum44. Osteocalcin appears to be involved in the mineralization process45. The proteoglycans, small proteins that are widely distributed in mammalian species, are also present in cementum46. Biochemical analysis of extracts of human cementum have identified chondroitin sulfate, dermatan sulfate and hyaluronic acid47. The enzyme alkaline phosphatase is believed to participate in cementum mineralization. In rat molars, the enzyme is heterogeneously distributed in the periodontal ligament, with the highest activity being found adjacent to alveolar bone and cementum. The enzyme activity adjacent to cellular intrinsic fiber cementum is higher than that to acellular extrinsic fiber cementum, and the thickness of the later correlates positively with the enzyme activity48. Several polypeptide growth factors with ability to promote proliferation and differentiation of putative cementoblasts are found in cementum matrix. These include BMP-2, -3 and –4, PDGF, α-and β-FGFs, TGFβ, PTH and IGF-13-4,49. While these molecules have been called growth factors, their role during tooth development does not appear to be related to proliferative activity. In fact, some TGFbs and PTH-related protein may have a role in regulating cell differentiation and subsequently mineralization49-50. It is important to know that many of these components are also present in bone; however, molecules unique to cementum have also been described. One of these is an IGF-1 isoform, referred to as cementum growth factor (CGF), now considered to be an insulin-like growth factor1-like molecule, with similar properties to those of IGF-1, but larger than IGF-1 in molecular size51. The second molecule is a collagenous protein referred to as cementum attachment protein (CAP). Antibodies to CAP immunostain only cementum and not other periodontal components or other tissues52. In bovine tooth germs, the CAP is expressed by cementoblasts, and in cementum its expression pattern is different from that of type I collagen53. CAP promotes the adhesion and spreading of mesenchymal cells; however, it promotes the adhesion of mineralized-tissue-forming cells, preferentially54-55. Cementum and Periodontal Regeneration
Although the extent of injury and the amount of lost tissue that must be filled are important determinants, whether a damaged tissue heals by regeneration or repair depends on two crucial factors: the availability of needed cell types and the presence or absence of the cues, and signals necessary to recruit and stimulate these cells. One major goal of regenerative periodontal therapy is new cementum formation and restoration of soft tissue attachment to the cementum. This process requires cementoblasts, and the origin of cementoblasts and the molecular factors regulating their recruitment and differentiation are not fully understood. In vivo animal models to evaluate cementogenesis during tooth development, the expression pattern of specific matrix molecules, and in vitro studies about the effects of cementum components on periodontal cells have provided important knowledge on how cementum components can regulate cementum regeneration3, 56-59. Although cementum formation in rodents differs from that in humans18, the observation of Liu et al (1997)60 indicates that periodontal ligament may be one source of cementoblasts progenitor cells in adult humans. These investigators demonstrated that a small proportion of clones of cells cultured from human periodontal ligament form cementum-like mineralized nodules in culture, and also produce cementum-specific markers61. Cementoblasts may also be derived from stem cells present in the periodontal ligament, gingiva, and alveolar bone, when the pool of available progenitors is likely to be reduced or absent; however, the molecules responsible for recruiting and differentiating these cells and the mechanisms involved remain to be better investigated3. A variety of chemotatic factors, adhesion molecules, growth factors, and matrix constituents participate together in the recruitment of cementoblasts progenitors, their expansion and differentiation. For example, cementum contains molecules that promote chemotatic migration, adhesion, proliferation and differentiation of some periodontal cells types better than others, and these molecules are not detectable in other periodontal structures51-52,55,61-63. Adhesion molecules that cause negative selection by excluding unwanted cells are also present in cementum54. Thus, the cementum microenvironment contains all the components necessary for cell recruitment, proliferation and differentiation, and molecules from the circulation are not necessary44. All these observations underscore the importance of restoring or providing the cementum microenvironment to initiate and promote new cementum formation. The integrity of cementum is altered by periodontal disease due to deposition of bacterial endotoxins64, and diseased cementum is removed during periodontal therapy65. Dentin not covered by cementum undergoes resorption. Root conditioning does not restore the original composition of cementum local environment, and instead expose molecules, especially type-I collagen, which has poor cell specificity65. The application of some growth factors is not likely to provide the complete repertoire of the needed molecules, since the concentration and type of growth factor on cementum change continuously during the healing process66. Similarly, while barrier membranes can facilitate population of the site by needed cells, they are not likely to provide the local environment for their differentiation67. Further, providing enamel proteins68, while likely to be conductive to early cementogenesis, may not provide appropriate environment for recruiting cementoblasts progenitors in adults and for their differentiation. Indeed, extracellular matrix components are expressed during periodontal healing67,69; however, whether all molecules are expressed in adequate temporal sequence is not known. All these factors together may explain why cementum regeneration is not always predictable for the available regenerative procedures. Recent in vivo studies using rat periodontal defect models, have shown that both BSP and OPN are expressed by cells linked to formation of mineralized tissues, while osteopontin is also expressed by cells within the newly formed periodontal ligament69. Future research directed at overexpressing or blocking expression of these molecules during periodontal wound healing should provide additional information required to establish the real function of these molecules in mineralized tissues and also determine the value of using such agents clinically for enhancing regeneration of periodontal tissues3. Future directions on Cementum research
The development of the cementum research in the cellular and molecular fields is promising, since several laboratories have reported successful isolation and propagation of cells (from both animal and human sources) exhibiting an apparent cementoblastic phenotype in vitro70-72. These cells have been shown, reproducibly, to form histologically proven cementum-like tissue following in vivo transplantation into immunodeficient mice44,72. One of the most important benefits of the establisment of such combined in vitro/in vivo models is that they will allow for elucidations of the relationship between osteoblasts and cementoblasts, besides providing more information regarding the specific mechanisms involved in maintenance of cementum structure and function in humans on the cellular and molecular level. Similarly, the in vitro/in vivo system can also be used for further elucidation of the modes of action of current available regenerative products, such as Emdogain, with apparent cementum-growth-promoting activities. This knowledge probably will allow for the development of strategies for regenerating cementum with cell-based therapies, by means of recruiting cells with cementoblastic potential, inducing their differentiation and excluding unwanted osteoblastic precursors. Furthermore, the use of these models allows for direct in vivo human experimentation, which may be particularly important, because cementum has been reported to differ between species in some aspects of its physiology9-11. In principle, cementogenic cells can be isolated from a relatively small specimen, expanded in culture, and subsequently transplanted to the same patient. Primary human cementoblasts can be efficiently grown from dissected fragments of healthy root cementum treated with colagenase: the culture conditions are fairly standard and it is possible to obtain large numbers (in the range of 108109) of committed cementogenic cells from a single tooth44,72. However, a significant drawback of this model is the requirement that healthy teeth be extracted for the cultures to be established, since it is still not clear whether cementogenic cells can be obtained, by this approach, of aged and/or diseased patients. These drawbacks will probably lead, in near future, to the investigation of sources other than cementum for cell isolation (i.e.: periodontal ligament, gingiva, bone marrow). Once the cementogenic cells are selected and the molecules critical for cementum regeneration are identified, the next step may be to develop an efficient and relatively simple delivery system to be used for periodontal regeneration. This probably may be one of the major problems, since cementum is a mineralized “interface” tissue, connecting mineralized dentin and non-mineralized fibrous periodontal ligament –what implies that a successful delivery system should somehow reconcile these two distinct environments. In this concern, molecules such as CAP show some promise60-62, due to its mineral-binding domain. Several integrins as well as their natural ligands (e.g. collagens, BSP, OPN) are known to be expressed by cementoblastic cells67, implying that they may be important regulators of cementogenesis. This also indicates that targeting integrin receptors may be beneficial for stimulating cementum regeneration. In summary, there is still much to study about cementum tissue regeneration via cell- and matrix-based therapies, but, without any doubt, recent developments in basic science indicate that these approaches are feasible and worth exploring. References
Figures source: Oral Histology Images. 2001 USC School of Dentistry and Health-Soft-Edu, at USC homepage www.usc.edu/dental. Copyright 2005 - Piracicaba Dental School - UNICAMP São Paulo - Brazil The following images related to this document are available:Photo images[os05003f3b.jpg] [os05003f1.jpg] [os05003f3a.jpg] [os05003f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}