 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
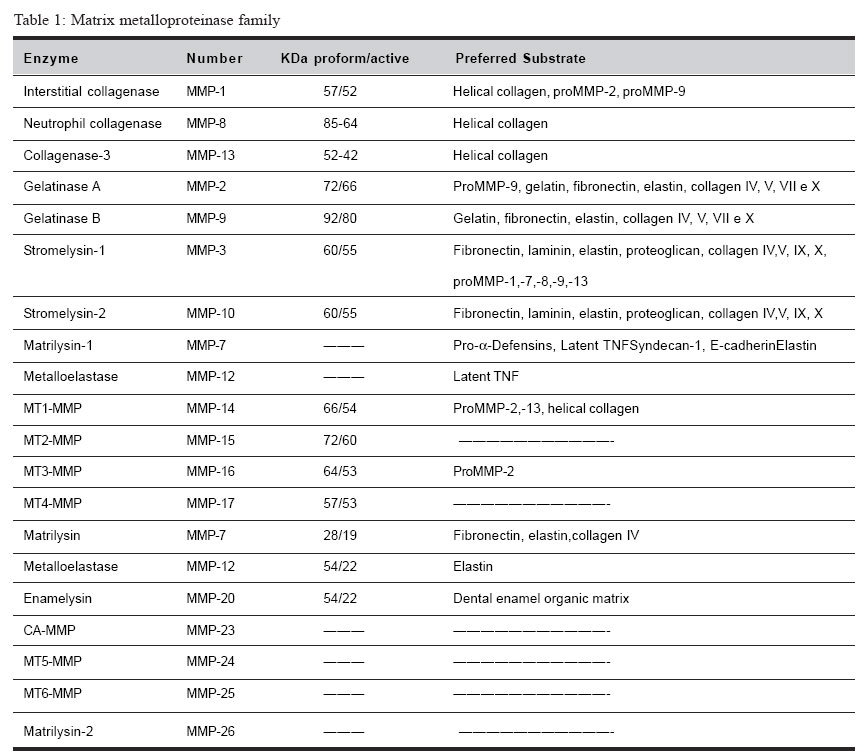
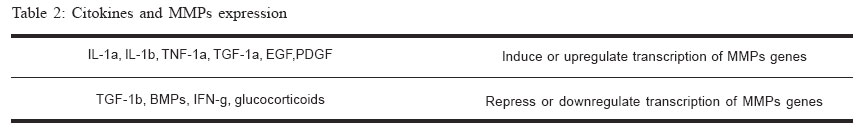
Brazilian Journal of Oral Sciences, Vol. 4, No. 15, Oct./Dec. 2005, pp. 884-890 Matrix metalloproteinases: the most important pathway involved with periodontal destruction De Souza, A.P.* Da Silva R.A.* Da Silva, M.A.D.* Catanzaro-Guimarães, S.A.** Line, S.R.P* *Department of Morphology, Piracicaba Dental School/University of Campinas -Piracicaba-SP/Brazil **Oral Biology Program - PRPPG/University of Sagrado Coração - Bauru-SP/Brazil Correspondence to: Ana Paula de Souza Oral Biology Program PRPPG-USC Rua Irmã Arminda 10-50 17011-160 - Bauru-SP - Brazil E-mail: anapaula_pardo@yahoo.com.brReceived for publication: September 02, 2005 Code Number: os05034 Abstract Periodontitis is an infectious disease estimated to occur in approximately a third of adults over the age of 35, being the major cause of adult tooth loss. The tissue destruction seems to be regulated by four major pathways. Plasminogen-dependent, phagocytic, osteoclastic and matrix metalloproteinase pathway. The matrix metalloproteinases (MMPs) pathway seems to be the most relevant in periodontal disease. The purpose of the current study was to review the roles of MMPs on periodontal disease, with emphasis on periodontal ligament and alveolar bone destruction. Particular attention is given on the mechanisms that control MMPs genes transcription, the regulation of protein activity, and the influence of MMP genes polymorphisms in inflammatory diseases. Key Words: MMP, cytokine, periodontitis, genes polymorphisms Introduction The Matrix Metalloproteinases Family Matrix metalloproteinases represent a family of dependent metal ions endopeptidases that are capable of degrading all extracellular matrix components, including several types of collagen and basement membrane components1-2. MMPs are metal-dependent since all members of this family have a zinc and a calcium-binding catalytic domain. They are secreted as inactive proenzymes (zymogens) and their activation occur in the tissue by cleavage of the N-terminal propeptide domain by other proteinases3. Presently, twenty-two MMPs have been identified in human tissues4-5. They are classified into different groups according to similarity in structure, gene encoded and substrate affinity5-6 (Table 17,8-9). Four major subgroups are directly related with the destruction of periodontium: collagenases, gelatinases, stromelysins and membrane-type MMPs (MT-MMPs). The collagenases are capable of degrade native type I collagen due to their helicase activity that attacks the triple helix of type I collagen. The released fibrils are then degraded by gelatinases or by lysosomal proteinases (phagocytic pathway). MMP-2 seems to have a helicase activity and to cleave fibrillar type I collagen10-11 . Although stromelysins (MMP-3) and membrane-type MMPs do not have helicase activity, these enzymes play a crucial role during activation of other MMPs, promoting the cleavage of the propeptide region, which that maintains enzyme latency6. The Importance of Matrix Metalloproteinases During Periodontal DiseaseInflammatory destruction of periodontal attachment apparatus is the hallmark of periodontal disease. The inflammatory reaction associated with periodontitis may damage the surrounding cells and connective tissue structures, including alveolar bone, causing tooth loss12-14 . During this event, the most important component of periodontium lost is the collagen type I that is found in the periodontal ligament and alveolar bone organic matrix. Four distinct pathways may be involved with this destruction: Plasminogen-dependent, phagocytic, osteoclastic and matrix metalloproteinase pathway1. A wide range of evidences has indicated that the most important pathway is the one which involves matrix metalloproteinases (MMPs)1. The most frequent cause of periodontitis is bacteria infection. The toxins, enzymes, and metabolites of bacteria present in the dental plaque play a key role in the initiation of the immune response that induces tissue destruction indirectly by activating host defense cells, which in turns produce and release inflammatory mediators. It stimulates the effectors of connective tissue breakdown. Lipopolysaccharides (LPS) derived from bacteria membrane have the capacity to activate junctional epithelial cells to release potent cytokines, such as interleukin-1, interleukin-8, tumor necrosis factor-a, prostaglandins, and proteases. Chemoattractant signals are started and transmigrating leukocytes and monocytes/ macrophages amplify local inflammation. In a second stage, cellular response is started. Resident ligament cells such as fibroblasts, macrophages, osteoblasts, keratinocytes, and endothelial cells are activated in response to stimulus, contributing with the synthesis of cytokines and MMPs. MMPs are present in both active and latent forms in chronically inflamed gingival tissues and gingival crevicular fluid. Active collagenase and gelatinase are found in the crevicular gingival fluid of patients with periodontitis in much larger amounts than in control subjects15. In contrast, high concentrations of the natural tissue inhibitor of MMPs (TIMPs) are found in the gingival crevicular fluid of healthy gingiva16. The area occupied by collagen fibers in gingival tissue specimens with periodontitis is significantly decreased, and the presence of MMP-1, MMP-2, and MMP9 is increased17-18. Matrix Metalloproteinases and Bone ResorptionThe role of MMPs during bone resorption is not totally clear but new insights suggest that MMPs also play a key role during this event. Osteoclasts are unable to attach to the bone surface if the mineralized bone matrix is covered by an osteoid layer19-21. Osteoid is composed by type I collagen, proteoglycans, glycoproteins, and native types IV and IX collagens. Osteoblast-derived collagenase (MMP-13) seems to be main responsible for degrading the nonmineralized osteoid layer covering bone surfaces, exposing the mineralized matrix to osteoclasts. MMP-13 (collagenase-3) is expressed in human breast carcinomas22, articular cartilage from arthritic patients23-26, osteoblasts, periosteal cells and fibroblasts during human fetal bone development27-28, and postnatally in bone remodeling28. MMP-13 is as efficient as MMP-1 and MMP-8 in the digestion of type I collagen26. Cleavage of collagen I by MMP-13 seems to be the initial step of the entire bone resorption process21. Subsequent, denatured collagen fragments are also degraded by gelatinases MMP-2 and MMP-9. MMP-9 is abundantly expressed in osteoclasts and it plays a distinct role to cathepsin K in the process of osteoclastic bone resorption. MMP-9 is cleaves acid insoluble type I collagen at 37°C and presents strong proteolytic activity against denatured type I collagen and type IV collagen29-31. Experimental evidences have suggested the importance of MMPs during bone resorption. Studies have demonstrated that MMP-3, MMP-9 and MMP-13 mRNA levels are increased when osteoblast cultures are stimulated by resorptive factors such as cytokines IL-1b and TNF-a, parathyroid hormone, and prostaglandin E2.21,32. Other direct evidence of the participation of MMPs in bone resorption is the fact that inhibition of MMPs by chemically modified tetracyclines can prevent bone loss33. The role of Citokines on MMP gene transcription At least three important regulatory mechanisms may control the action of MMPs on the components of extracellular matrix: regulation of transcription levels, activation of the proenzyme into the extracellular matrix (enclosing the plasmin-dependent or MMP-dependent pathway), and the inhibition of MMPs by their tissue inhibitors of MMPs (TIMPs). Control of transcription has been considered the key step in the regulation of MMPs activity. However, the control of protein activity is of key importance. MMPs activity is controlled by endogenous activators, inhibitors and factors that influence MMPs secretion, cell surface localization, and MMPs degradation and clearance. MMP genes are expressed when required in a physiological extracellular matrix remodelation or during pathological tissue destruction34-36. MMP-2 (gelatinase A) represents an exception. It is secreted constitutively and low rates of MMP2 are found in normal tissues. So, MMP-2 activity is mainly controlled by enzyme activation37 and post-transcriptional mRNA stabilization38. The expression of several MMPs can be up- or downregulated by extracellular matrix proteins, cell stress, phorbol esters, and changes in cell shape39. MMP expression can be still induced by its own substrate and specifically repressed, once cleaved the substrate it is no longer needed. For instance, type I collagen acts as a ligand for discoidin domain-containing receptor-like tyrosine kinases. This enzyme induces MMP-1 expression when activated by intact collagen and become inactive after binding MMP-1 cleaved collagen40-41. The promoters of many MMP genes (including MMP-1,-3,7,-9,-10,-12 and –13) contain an AP-1 consensus element located approximately at -70. Although necessary, the presence of the AP-1 binding site is not sufficient to activate the transcription of MMP genes. The transcription is achieved by a concerted action of the AP-1 site with other regulatory sites such as the ones for Fos and Jun and/or the Ets family of transcription factors. The promoter of MMP genes is susceptible to activation by several citokynes, growth factors, and the extracellular matrix metalloproteinase inducer (EMMPRIN)35,42 (Table 21). Normal periodontal cells (fibroblasts, keratinocytes, osteoblasts, endothelial cells, macrophages) and inflammatory cells can be stimulated by citokines. A same signaling molecule may regulate the expression of several MMPs. TGF-b suppresses the transcription of the MMP-1 and MMP-3 genes, but induces the expression of MMP-13 (collagenase-3)43. On periodontal fibroblasts IL-1b and TNF-1a can upregulates collagenase (MMP-1), gelatinase-A (MMP-2) and stromelysin1 (MMP-3) synthesis1. IL-1b and TNF-1a have the same effect on of osteoblast collagenase (MMP-13) production44-45. IL-6 upregulates osteoblasts gelatinase-A (MMP-2)11. On the other hand IL-1b appears not to affect keratinocytes. TGF-1b represses the transcription of most MMPs but upregulates MMP-21. IL-8 recruits neutrophils to inflammatory sites and it is able to promote neutrophil degranulation that releases large amounts of gelatinase-B (MMP-9) into periodontitis sites46. Recent studies have revealed the existence of MMP substrate binding sites termed exocites located outside the catalytic domain that are able to cleave noncollagenous substrates such as chemokynes and some citokynes47. This cleavage results in small peptides that may be more potent activators than the whole fragment. IL-8 provokes the releasing of MMP-9 of neutrophil granules and is a potential target of these enzymes46. Proteolysis of non-matrix proteins such as cytokines, receptors and adhesion molecules is also being increasingly recognized as an equally important aspect of chronic inflammatory disease. Proteinases of the MMP family take an important part in the processing of these bioactive molecules and degradative events. Polymorphisms in Matrix Metalloproteinase Promoter RegionsThe cis-regulatory DNA of many genes is organized into independent modules that regulate the transcription of specific tissues at specific times48. The regulatory modules are usually constituted of multiple binding sites for transcription factors that can be located near the transcription start site, within introns or thousand of base pairs away from the regulated gene49. Mutations within individual modules can enhance or repress gene transcription in a tissue specific manner. Accordingly, gene polymorphisms in cis-regulatory sequences can interfere with the transcriptional regulation of genes. The most important region for transcriptional regulation is known as “promoter”. These regions are usually located upstream from transcription start site and contain sequences for the binding of critical transcriptional factors. Promoters usually work in concert with other cis-acting sites (enhancers and silencers) to direct the levels of transcription of a certain gene. Several DNA polymorphisms have been found in the promoter regions of several MMPs. Polymorphisms represent natural sequence variants (alleles), which may occur with more than one form, having a frequency greater than 1% in a human population. Basal and inducible levels of MMP gene expression can be influenced by these genetic variations that may influence the development or progression of several diseases, including periodontal disease. An insertion/deletion of a guanine at position –1607 of human MMP-1 gene creates two different alleles, one having a single guanine (1G) and the other having two guanine (2G)50. The 2G allele together with an adjacent adenosine creates a core binding site (5‘-GGA-3‘) for Ets family of transcription factors immediately adjacent to an AP-1 site, causing a 37-fold increase in transcription activity36,42,50. Tumors bearing the 2G allele can secrete higher levels of MMP-1, and the presence of this allele was associated with the development of ovarian cancer51. Moreover, an unusually large proportion of metastatic melanomas with loss of heterozygosity at this site retain the high expressing 2G allele52. A case-control study performed in 87 non-smoking Caucasian subjects showed a positive association between the 2G allele and the severity of chronic periodontal disease53. This same allele was also associated with early dental-implants failure54. Another study performed in a Japanese population has found no association between this MMP-1 polymorphism and periodontitis55. These results may suggest that genetic variation in different races may promote differences during pathological process. Two functionally important genetic polymorphisms have been detected in the promoter of the MMP-9 gene. One represents a SNP at position -1562 and the other a (CA)n microsatellite repeat at position -90. The SNP is a C to T substitution that increases the transcriptional activity56. The C-1562T polymorphism has been associated with of coronary atherosclerosis57. The (CA)n is a multi-allelic microsatellite polymorphism where the most common form is the (CA)14. The 14CA repeat has only 50% of the transcriptional activity of MMP-9 promoter comparing with the 21CA repeat allele58. Associations between the (CA)n repeats and abdominal aortic aneurysm and intracranial aneurysm have been reported, but the data are contradictory59-62. A case control study with 100 non-smoking subjects failed to show a positive association between the MMP-9 C-1562T polymorphism and the susceptibility/severity to chronic periodontitis63. A 5A/6A functional polymorphism at position in the -1171 of the MMP-3 (stromelysin-3) gene promoter has been associated with atherosclerosis in a number of genetic epidemiological studies64-65. The frequency of the 5A allele is significantly higher in affected individuals than in control subjects and the risk of acute myocardial infarction in individuals carrying one or two copies of the 5A allele was estimated to be 2.25 fold36,66. MMP-3 is capable to degrade a wide range of extracellular matrix proteins, promoting the cleavage of atherosclerotic plaque. These data indicate that the 5A/6A repeats are of potential interest in studies of DNA polymorphism-periodontal disease association. The evidences presented suggest that genetic polymorphisms in the MMPs genes are likely to be related to a wide range of diseases that are characterized by extracellular matrix degradation, including periodontal disease. Matrix metalloproteinases, TIMP Complexe, and Synthetic Inhibitors of MMPsThe activity of MMPs proteinases in the extracellular matrix (ECM) is regulated by specific inhibitors of MMPs known as tissue inhibitors of metalloproteinases (TIMPs)7. Presently, the TIMP family consists of four members (TIMP1, -2, -3, -4). TIMPs inhibit the proteolytic activity of activated MMPs by forming a 1:1 stochiometric inhibitory complex with the enzyme. Disturbances in the MMP/TIMP systems have been implicated in the etiology of diseases67 such as arthritis, pulmonary emphysema, atherosclerosis and periodontitis, where a loss of ECM is a major feature68. The search for a synthetic inhibitor of MMP activity has been the aim of pharmaceutical companies since 1970, when ICI (now AstraZeneca) began the first clinical trial using MMPs inhibitors for the treatment of arthrtitis69-70. A number of MMPs inhibitors have been synthesized but the major drawback of these molecules are the side-effects. Musculoskeletal syndrome has been reported as the most common side-effect in clinical studies using MMP inhibitors. Events such as joint pain, stiffness, edema, skin discoloration, and reduced mobility are observed. In arthritis, the inhibition of MMPs surprisingly exacerbates rather than alleviates the disease71. Presently, Periostat® (doxycycline hyclate, CollaGenex Pharmaceuticals Inc.) is the only collagenase inhibitor approved by the FDA and clinically employed. Doxicycline is a chemically modified tetracycline without anti-bacterial activity. This substance does not promote musculoskeletal syndrome side-effect and is capable of inhibiting collagenase activity. Studies have demonstrated that low dose doxycycline used as an adjunct to scaling and root planning (SRP) significantly improves clinical attachment and reduces pocket depth, slowing the progression of periodontitis when compared with SRP alone72. The benefits were achieved after three months of treatment, and appear to be maintained during a 3-mont post-treatment period. However, the therapies are not totally successful. The attachment level gains in the deepest sites are modest in these studies (~0.7 mm). The gains in deeper sites treated with systemic tetracyclines and SRP are twice bigger than gains achieved using MMP inhibitor70. In conclusion, MMPs play an important role during the process of periodontitis since they are responsible for the degradation of extracellular matrix components. In recent years the use of molecular and cellular biology methodologies has greatly contributed to the understanding of the basic mechanisms that determine the onset and progression of periodontal diseases. It is likely that in a recent future they may also help in the prognosis and in the prediction of disease progression. This will facilitate the identification of individuals at high risk, allowing the planning of individualized approaches of periodontal therapy. AcknowledgementThe research work of A.P. Souza has been supported by Fapesp grants 99/11866-0 and 03/10422-8. References
Copyright 2005 - Piracicaba Dental School - UNICAMP São Paulo - Brazil |

{kind=link}
{kind=link}