 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
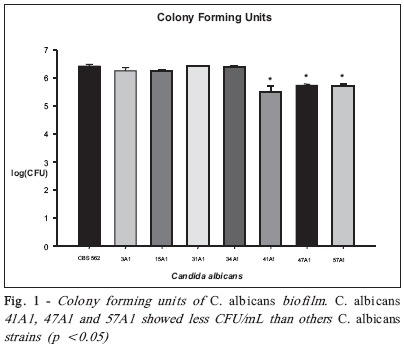
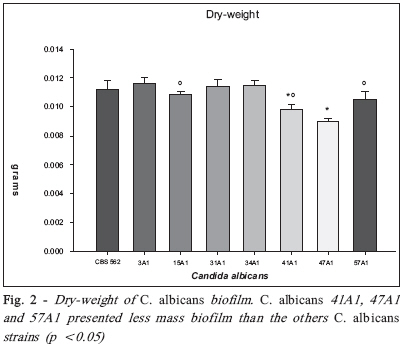
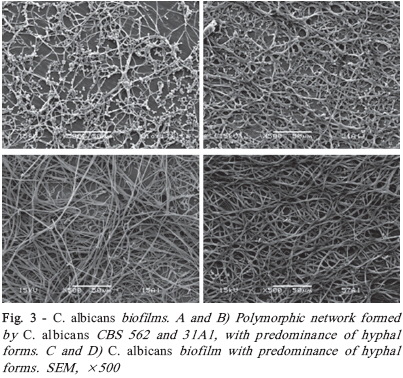
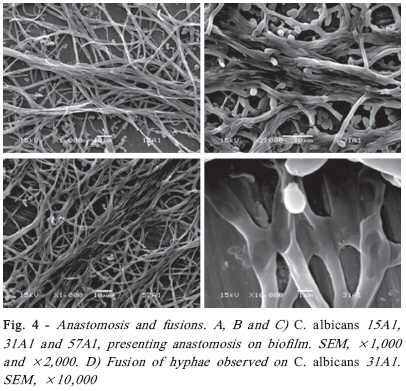
Brazilian Journal Oral Sciences, Vol. 7, No. 27, Oct/Dec, 2008, pp. 1678-1681 Evidence of Heterokaryon compatibility on Candida albicans biofilm Gustavo Alberto Obando-Pereda1; Paula Cristina Anibal1; Vivian Fernandes Furletti1; José Francisco Hofling2 1DDS, MS, Graduate Student, 2B, MS, PhD, Professor Department of Microbiology and Immunology, Piracicaba Dental Schoolf, University of Campinas, Piracicaba, SP, Brazil Received for publication: July 30, 2008 Accepted: September 30, 2008 Code Number: os08043 Abstract Aims: Candida species, especially Candida albicans, are frequently found associated with biomaterials and immunosuppressed patients,
and have been described as the most virulent yeasts in human fungi diseases. These yeasts have recently been isolated from periodontal
pockets, revealing the penetration of hyphae into the periodontal connective tissue. Keywords: C. albicans, biofilm, heterokaryon compatibility. Introduction Candida yeasts are opportunistic pathogens that cause diseases in hosts that are compromised by underlying local or systemic pathological processes1. Candidiosis is the most common fungal infection in humans2 . Candida albicans is an aerobic/anaerobic commensal that can be cultured from the oral cavity of young adult3-4. In the oral cavity, yeasts commonly colonize the tongue, palate, and oral mucosa3 , and may occur in subgingival plaque of adults with severe periodontitis5. It is estimated that around 75% of healthy people care members of the genus Candida in skin and mucosa, and 36% of this population have shown evidence of oral candidiasis6-7. In healthy carriers, various local and/ or systemic general predisposing factors give Candida species the capacity to invade different mucosal tissues, making them opportunistic pathogens6-7 . C. albicans is the main specie responsible for the majority of oral mycotic infections, although other members of this genus may be involved6,8. Like other pathogens, virulence in C. albicans includes host recognition (adhesion, co-aggregation to host cells and biofilm development), several degrading enzymes, hyphal formation and phenotypic switching9-10 . When fungal colonies grow contiguously, the hyphae may fuse (anastomosis) to form cells with nuclei and cytoplasm from both parent colonies (heterokaryon)11. The biological significance of this phenomena explain the horizontal transfer of genes, acquisition of new biological characteristics, homeostasis and cellular communication12 , but on other hand, this phenomena works like a major histocompatibility complex that preserves the integrity of the organism, limit the heterokaryon formation (heterokaryon incompatibility) preventing the transfer of infectious cytoplasmic elements, virus transposons and debilitated organelles13. These genes are called het (for heterokaryon formation) and have been extensively studied in Podospora anserine. One of these genes, called het-s, has two alleles, het-s and het-S. Strains with the same allele can undergo hyphal anastomosis to form heterokaryons, but when het-s and het-S strains grow together, heterokaryon incompatibility is observed11. In oral biofilms, microbial communities consist by more than 700 different microbial species14. In subgingival biofilms have been described many bacterial complex and yeasts belonging to Candida species5,15. Moreover, several studies have reported virulence factors for C. albicans, such as like adhesion, proteinase secretion, hyphal formation, phenotype switching, biofilm growth and antifungal resistance10. Considering that biofilm is a vital and natural structure for microorganisms to exchange nutrients and genetic information and that periodontal C. albicans is a common yeast in periodontal biofilms associated with periodontal microbial complexes, and given that heterokaryon compatibility is an phenomena that allows for nutrient and genetic material interchange in fungi, the aim of this study was to investigate the heterokaryon formation of periodontal C. albicans strains in biofilms. Material and Methods Yeast Preparation and Adherence Assay Procedures Samples of C. albicans CBS-562, and the periodontal isolates 3 A1, 15 A1, 31 A1, 34 A1, 41 A1, 47 A1 and 57 A1, which were positive to phospolipases and aspartyl proteinases16, were obtained from the culture collection of the Microbiology and Immunology Laboratory of Piracicaba Dental School, Brazil. Candida species were reactivated from their original cultures (Sabouraud agar, SDA) at 37ºC in 5 mL of Brain Heart Infusion (BHI) supplemented with 0.5% of yeast extract granulated (BHIY) and incubated for 24 h at 37ºC in an aerobic atmosphere. The species were harvested at 2,400 xg for 10 min, suspended in 10 mL of NaCl 0.89% buffered with sodium phosphate 0.1M, pH 7 (PBS, phosphate buffer solution), and agitated for 15 s. After this procedure, the cell concentration was measured spectrophotometrically at 520nm ensuring an OD 800 equivalent to 1 x 106 CFU/mL17 . Titanium discs (Conexão Sistema de Prótese Ltda., São Paulo, SP, Brazil) were placed in 24-well tissue culture plates (Nunclon; Nunc, Roskilde, Denmark) and 80 μL of standard yeast suspension were dropped on the discs. After 180 min of cell adherence, the non-adhered microorganisms were removed by PBS washing18. Next, the titanium discs were placed on a new tissue culture plaque and submerged in 2 mL of BHI for incubation at 37ºC in aerobic atmosphere. After 48 h, the discs were washed with PBS and sonicated. Candida cells were plated on SDA for counting of CFU/mL19-20. Dry-weight was carried out according to Koo et al. (2003)21. Briefly, the removed biofilms were subjected to sonication and the homogenized suspension was used to dry weight determination. Three volumes of cold ethanol (-20°C) were added to 5 mL of the cell suspension and the precipitate was collected (10,000 xg for 10 min, 4°C). The supernatant was discarded, and the cell pellet was washed twice with cold ethanol, and then lyophilized and weighed. Scanning Electron Microscopy Some samples were fixed in 2.5% glutaraldehyde, 0.15M PBS (v/v) for 1 h at room temperature. Next, the discs were treated with 1% osmium trioxide for 1 h and washed three times with 3 mL of distilled water. The discs were hydrated in an increasing ethanol series (50% to 100%), critical-point dried, sputter-coated with gold and examined under SEM22. This test was done in triplicate for each sample. The anastomosis was identified like a many contiguous hyphal growing fused between them, and the fusion, like a hyphal fusion between two cells. Statistical analyses All statistical tests were performed by one-way ANOVA and Bonferroni’s correction at 5% significance level using Sigmastat 3.0 Demo system software (SPSS Inc., Chicago, IL, USA) and the graphs were done using Sigmaplot v.8.02 Demo system software (SPSS Inc.). Results C. albicans samples showed biofilm development on the titanium discs. The 41 A1, 47 A1 and 57 A1 samples produced a smaller number of CFU/mL (Figure 1) and less biofilm mass (Figure 2), p <0.05. SEM analysis of Candida biofilms showed a consisted polymorphic cell network, as reported by Al-Fattani and Douglas (2004)23, with predominance of hyphal morphology (Figure 3). All strains showed the presence of hyphal anastomosis, but 31A1, 57A1 and 15A1 exhibited more anastomoses and fusions between hyphae, proving the heterokaryon compatibility of C. albicans strains (Figure 4). Discussion The development of biofilm by C. albicans has been fully described22,24. This biofilm is characterized as a network of polymorphic cells, yeasts and hyphae with extracellular matrix production20,22,24, but few studies have shown anastomosis between hyphae and fusions occurring in the biofilm25-26. Many virulence factors described in the literature, such as adhesion, several degrading enzymes, hyphal formation and phenotype switching, confer a virulent potential to C. albicans strains, isolated mainly from immunosuppressed patients9-10. Biofilm is a vital, important and natural structure where microorganisms interchange genetic material and nutrients and, where the strains acquire resistance to antimicrobial agents and new virulent characteristics19 . The heterokaryon compatibility observed in biofilms models supports the evidence that these strains can fuse with others C. albicans clones or yeast species to interchange genetic material, nutrients and other elements providing new virulence characteristics. The importance of studies investigating heterokaryon compatibility as a virulent factor of C. albicans relies on fact that het-s/het-S strains can interchange many elements around the biofilm network, increasing C. albicans resistance to antimicrobial agents and its virulence, changing from harmless commensal yeasts to fatal human pathogens. In periodontal diseases, bacteria are continuously interchanging genetic material in biofilms. Yeasts can capture and integrate this genetic material in their genomes and exchange it with other yeasts by the heterokaryon compatibility phenomenon, creating a pathogenic entity in periodontal pockets. In conclusion, periodontal C. albicans strains present heterokaryon compatibility. Further research is needed to determine the importance of heterokaryon compatibility as a virulent factor of C. albicans. Acknowledgements This work was supported by the São Paulo State Research Foundation (Grant #2005/55981-0) and the Brazilian National Council for Scientific and Technological Development (Grant #132518/2005-8). References
© Copyright 2008 - Piracicaba Dental School - UNICAMP São Paulo - Brazil The following images related to this document are available:Photo images[os08043f3.jpg] [os08043f4.jpg] [os08043f2.jpg] [os08043f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}