 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
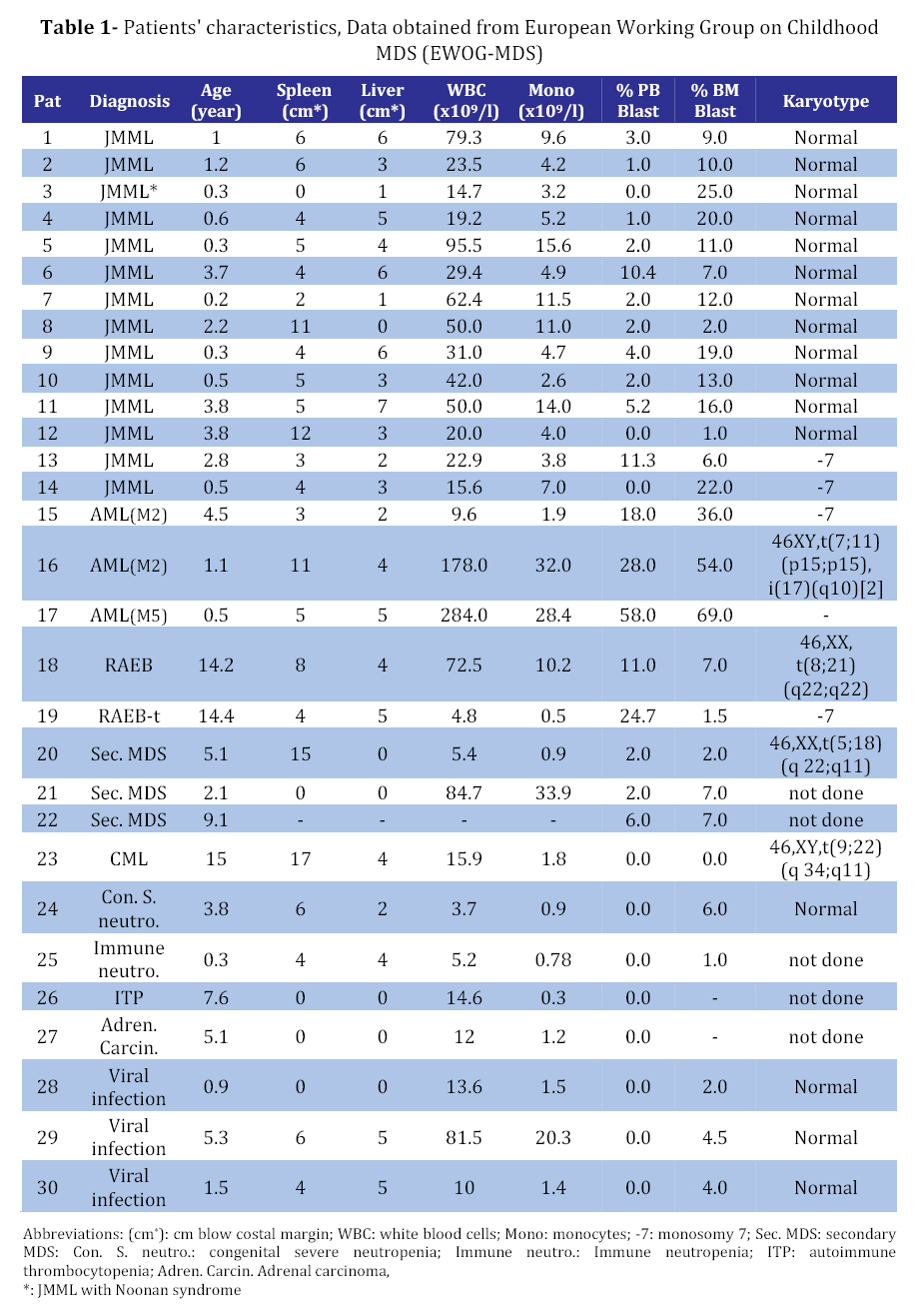
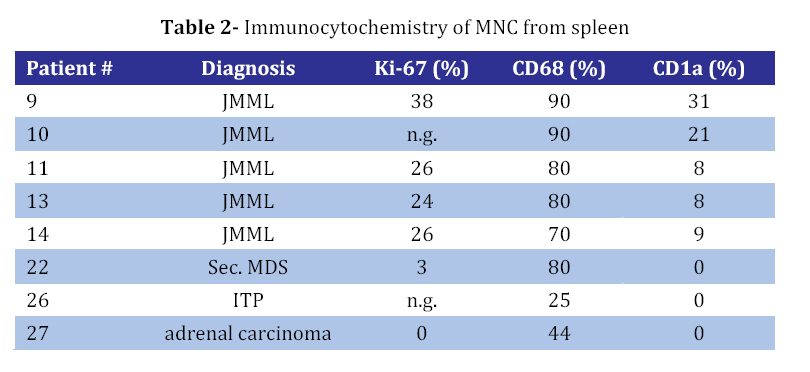
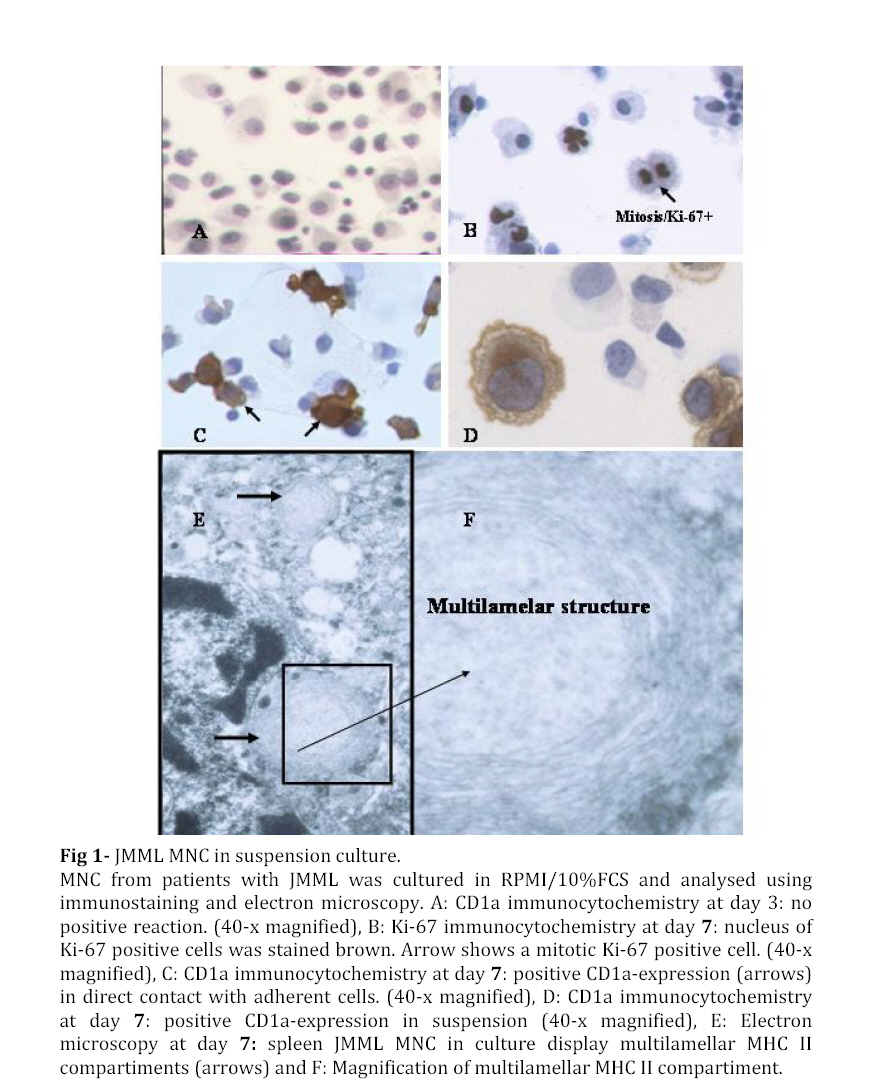
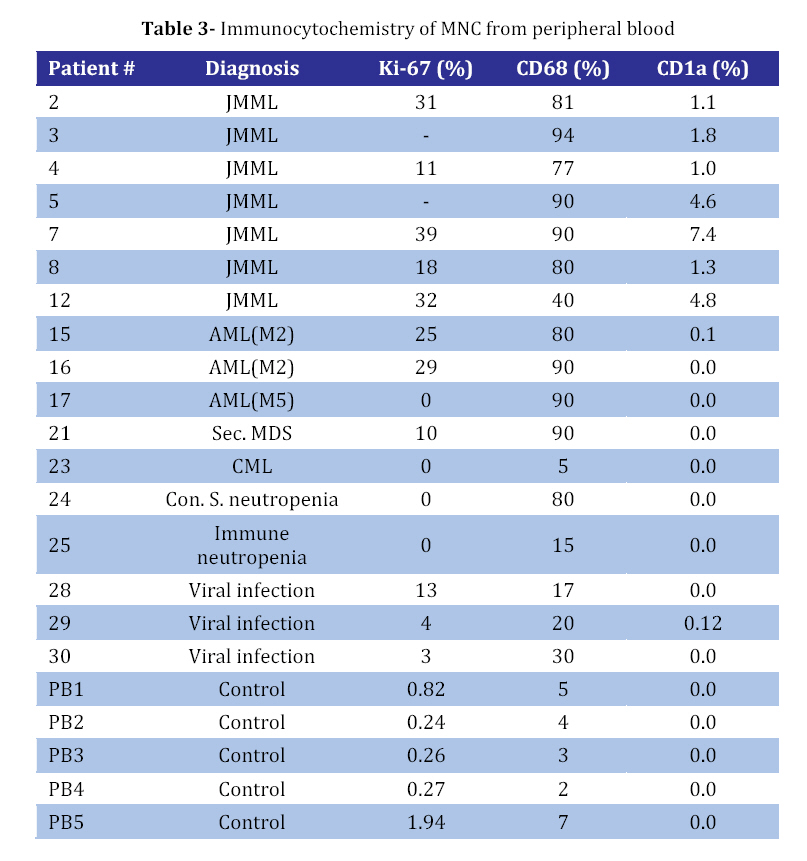
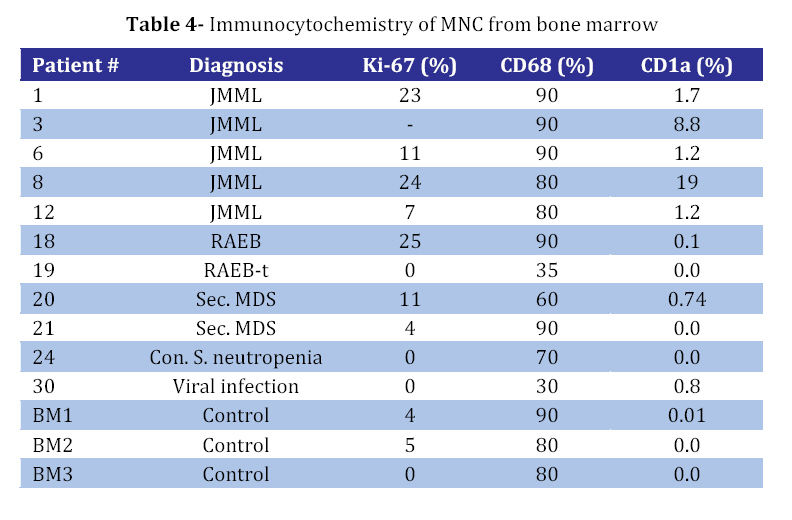
Iranian Journal of Pediatrics, Vol. 18, No. 3, Sept, 2008, pp. 213-221 Juvenile Myelomonocytic Leukemia (JMML) Cells Spontaneously Differentiate into Dendritic-cell Like Populations Parviz Shayan*1,2, DVM, PhD 1. Department of Pathobiology, University of Tehran, Tehran, IR Iran Code Number: pe08034 Abstracts Objective:Juvenile myelomonocytic leukemia (JMML) is a rare myelodysplastic/ myeloproliferative malignancy of early childhood, characterized by monocytosis, hepatosplenomegaly and an aggressive clinical course. Key Words: Juvenile myelomonocytic leukemia; CD1a; Dendritic cells; Suspension cell culture; Electron microscopy; Immunostaining Introduction Juvenile myelomonocytic leukemia (JMML) is a rare clonal malignancy of infancy, which accounts for approximately 2% of hematological neoplasias in childhood[1]. According to the WHO classification JMML is considered a bridging entity within the category of myelodysplastic/ myeloproliferative disorders [1,2]. Marked hepatosplenomegaly, lymphadenopathy and tumor cell infiltration of various organs such as lungs, intestine, liver, spleen and skin are common indicative features. They determine the clinical picture with organ dysfunction, infection and bleeding, and contribute to a high mortality[3,4]. Approximately 65% of patients with JMML have a normal karyotype whereas 25% display a monosomy 7, and 10% have complex chromosomal abnormalities[5]. Although there are no specific diagnostic chromosomal markers, in up to 75% of JMML patients mutations or other genetic abnormalities can be found in three distinct genes, namely the RAS (NRAS, KRAS2), neurofibromatosis type 1 (NF1), and PTPN11 gene. They are all positioned in the granulocyte-macrophage colony-stimulating-factor (GM-CSF)/RAS pathway[6]. Dysregulation of the signal transduction cascade results in a selective hypersensitivity of myeloid progenitor cellsto GM-CSF in vitro[7]: JMML cells have the capability to grow in semisolid culture without exogenous growth factor supplementation. This ‘spontaneous growth’ seems to depend on the presence of adherent macrophages, which presumably secrete cytokines and growth factors such as GM-CSF and tumor necrosis factor alpha (TNF-α), and herewith exert paracrine and/or autocrine stimulation of colony growth, proliferation and differentiation of the JMML cells[8,9,10].Here, we present a novel liquid suspension culture system for JMML, with easily accessible cells for various studies. We describe morphological, immunological and electron microscopic characteristics of mononuclear cells (MNC) from PB-, BM- and spleen of 14 patients with JMML and 24 control samples cultured for a minimum of seven days. Subjects & Methods Patients and Controls: Table 1 depicts the clinical characteristics of 14 patients with JMML, nine patients with other hematopoietic neoplasias (AML [n=3], MDS [n=5], CML [n=1]), one patient with adrenocortical carcinoma, and six patients with non-malignant disorders: congenital severe neutropenia [n=1], autoimmune thrombocytopenia [ITP, n=1], autoimmune neutropenia [n=1], viral infections [n=3]). In addition, eight healthy volunteers were studied. PB, BM or spleen tissues were obtained at the time of diagnosis with informed consent from all individuals or their parents conforming to our institutional review board guidelines. Isolation of Mononuclear Cells: Spleens were utilized as source of MNC since the resected organ provides abundant infiltrating leukemic cells, whereas PB or BM are scarce due to the young age of JMML patients and the low incidence of the disease. Spleens (n=8) were cut into small pieces and placed in phosphate-buffered saline (PBS). Cells were extracted by mechanical scraping and filtering (70 µm strainer). MNC from this cell suspension as well as from PB or BM were isolated by discontinuous density gradient centrifugation. Briefly, cells were layered on Ficoll- Paque (Pharmacia, Uppsala, Sweden) and centrifuged for 45 min at 18°C and 400 g without break. The MNC-containing interphase was transferred into IMDM/2% fetal calf serum (FCS) washed and either used directly for culture or stored at a concentration of 2x107 cells/mL FCS/DMSO in liquid nitrogen at -120°C. Suspension Cell Culture: Cryopreserved cells were thawed in a 37°C water bath and suspended in 20 mL RPMI/10% FCS/1% penicillin-streptomycin. 0.5 mg/mL DNase I (Desoxyribonuclease I, Sigma, Steinheim, Germany) and Liquemin (20 IE/mL) were added to avoid agglutination of cell debris. After washing MNC were cultured at a concentration of 1x106/mL in RPMI/10% FCS/1% penicillin/streptomyxin for 7 days at 37°C in an atmosphere containing 5% CO2. No supplementary growth factors were added. For morphology and immunocytochemistry studies 5x104 cells from the suspension were fixed to a cytospin slide by centrifugation at 800 rpm for 10 minutes. For separate analysis of the adherent cells in the culture cover slips were placed on the bottom of the plate (MBST, Iran) and removed for staining at distinct time points. Immunocytochemistry: Cells were stained with immunoperoxidase systems (DAKO StreptABComplex/HRP duett, mouse/rabbit) using murine anti- human-CD1a (clone NA1/34), -CD68 (clone KP1) and Ki-67 (code No. A0047) antibodies (DAKO Hamburg, Germany). After 10 minutes fixation in 4% formaldehyde cytospin slides were washed in TBS and incubated in 0.6% H2O2/methanol to eliminate residual peroxidase activity and in 5% FCS/TBS to block unspecific protein binding for 10 minutes, respectively. The primary anti-human-CD1a antibody was used at a dilution of 1:100, incubation duration was 1h, followed by addition of a Biotin goat anti-mouse-immunoglobulin antibody and a Strepatvidin/biotinylated (horseradish) peroxidase complex (StreptAB-Komplex/HRP) for signal intensification. A chromogen-substrate-solution of H2O2 and diaminobenzidin was used to visualize the signal. Slides were counterstained for 90 s in hematoxylin. As negative control, the slides were stained with mouse serum as the first antibody. The percentage of cells positive for Ki-67, CD1a and CD68 in at least 1000 cells. For control samples with low number of CD1a positive cells the number of CD1a positive cells on the slide was counted, while the total number of cells on the slides was estimated. Electron microscopy (EM): At day 7 1x107 cells from patient #9 were harvested from the suspension, fixed with 3% glutaraldehyde for 4h and post-fixed with 1% osmium tetroxide OsO4. After washing specimens were dehydrated in graded ethanol and embedded in Epoxy medium (1,2-epoxypropane, followed by 1,2-epoxypropan-epon-mix (1:1)). Ultra-thin sections (50nm) were placed on copper grids and stained with 2% uranyl acetate and 2% lead citrate. Examination followed using a Zeiss EM 10 TEM. Findings Generation of CD1a positive cells in suspension culture of JMML cells culture with MNC from splenic tissue of JMML patients were cultured in suspension in media containing 10% fetal calf serum without exogenous growth factors added[11,12]. Cells proliferated as indicated by frequent mitosis, increase in cell number and expression of the proliferation marker Ki67. At day 7 of culture, 24%-38% (median 25%) of non-adherent cells were Ki67 positive (Table 2, Fig 1B) and greater than 70% expressed the monocyte/ macrophage marker CD68 (Table 2). After day 5 of culture and with a maximum on day 7, large cells with irregular-shaped convoluted nuclei and widespread cytoplasmic projections were consistently noted. By immunophenotyping, these cells were characterized as CD1a positive (Fig 1D). It is to denote that the cultured cells are until day 5, CD1a negative (Fig 1A). The percentage of CD1a positive cells in suspension ranged between 8% and 31% (Table 2). On the cover slips CD1a positive cells were located in direct neighborhood and often with cell-to-cell contact with macrophages (Fig 1C). Control cultures with splenic MNC from 3 patients with secondary MDS, ITP or adrenocortical carcinoma contained up to 80% CD68 positive cells on day 7, but only scarcely Ki67 and no CD1a positive cells (Table 3). Culture with mononuclear cells from bone marrow and peripheral blood: Under the same culture condition as described above, generation of CD1a positive cells was also noted in JMML specimens from BM and PB, with 1.2-19% and 1.0-7.4% of all non-adherent cells on day 7 being CD1a positive, respectively (Table 3and 4). In other disease states including myeloid neoplasia or healthy controls CD1a positive cells were only seen in 6 of 24 cases, and maximum number did not exceed 0.7%. In all JMML samples and in some samples of AML and MDS patients cell proliferation with an increased percentage of Ki68 positive cells was observed. Electron microscopy of JMML-CD1a-positive cells: For further characterization of their ultra structure CD1a-positive cells were analyzed by electron microscopy. The cells exhibited pronounced membrane protrusions, lobulated nuclei and contained atypical granular multilamellar organelles. By day 7, these atypical granules appeared as onion-skin like structures in a substantial proportion of the CD1a-positive cells (Fig 1E and F). They have been described as multilamellar MHC II compartiments in DC. However, Birbeck granules, the characteristic cytoplasmic structures of DC of the Langerhans type were not detectable. Discussion To gain a better understanding of the mechanisms involved in the proliferation and differentiation of JMML cells in vitro, we established a culture system of JMML MNC without exogenous growth factor supplementation. As expected, cultures were characterized by marked adherence of cells and proliferative activity with frequent mitosis and a high Ki67 index. Remarkably, after day 5 of culture, all JMML specimens featured differentiation into CD1a-positive cells. These CD1a-positive cells were detectable in the adherent cell population, and accounted for up to 31%, 19% and 7.4% of cells in suspension of JMML spleens, bone marrow and peripheral blood, respectively. In contrast, only an occasional sample from specimens of other disease states or healthy volunteers displayed small numbers of CD1a-positive cells (≤0.8%). DC comprise a heterogeneous population of antigen presenting cells (APC), expressing CD1a, a non-polymorphic class I molecule, and MHC class I and II molecules[13]. Immature DCs have abundant complex vesicles, with multivesicular and multilamellar morphologies, consisting of large amounts of internal membrane. Different culture methods have allowed the in vitro generation of DC from CD34+ precursors, usually by the sequential use of different cytokine-combinations such as GM-CSF, IL-4 and/or TNF-alpha or SCF+IL3+IL6 followed by GM-CSF+IL4[14]. Likewise, culture assays from monocytes have been described, using GM-CSF, IL-4 and TNF-alpha[15]. Whereas Langerhans cells (LC), the immature DC of the epidermis, typically display CD1a and so-called Birbeck granules, in vitro-generated DC from CD14- positive monocytes express CD1a, but do not have typical Birbeck granules. However, according to ultrastructural studies, they display atypical multivesicular compartiments and phagocytic particles. Generally, membrane structures are rather interrupted, their width is larger, and they never seem to form from the plasma membrane[13,16]. Despite the lack of additional growth factor supplementation in our culture system, we observed the spontaneous differentiation into populations with DC- like features, such as the pronounced expression of CD1a and CD80 (not shown), the latter serving as a co-stimulatory molecule in the antigen-presenting function of DCs. Further evidence for their DC-character was obtained by electron microcopy showing abundant multilamellar/ multivesicular granules, which resemble major histocompatibility complex (MHC) class II compartiments (MCIIC)[14]. As indicated by FICTION analysis, these CD-like CD1a-positive cells belonged to the JMML clone[11]. Presumably, the ‘sponataneous’ differentiation of JMML MNC into DC-like cells is mediated via para-/autocrine mechanisms, such as the IL-4, IL-1, GM-CSF and/or TNF-alpha secretion[17]. Accordingly, addition of anti-TNF-alpha and GM-CSF monoclonal antibodies caused a marked reduction of CD1a-positive cells in JMML cultures and transfer of JMML-supernatant to healthy MNC cultures revealed a slight increase in CD1a-expression[12]. These observations allow the speculation of an existing growth factor secretion of the cultured JMML-DCs, in combination with a hypothetical hypersensitivity of these JMML-populations in vitro (similar to the known GM- CSF hypersensitivity). DCs in general have been described in association with a variety of neoplasias, however; rather exert immuno-protective functions by presenting tumor antigens to T-cells. Unlike ‘healthy’ DCs, an impairment or dysfunction of immunocompetence of JMML-derived DCs seems likely and will have to be addressed in further studies. There is evidence, that these cells lack the production of the T-cell/immunostimulatory cytokine IL12 but rather, seem to secrete the immunosuppressive IL10, which inhibits the differentiation and function of the APC[16] and therefore seem to have defective capacity to prime naive T cells[17]. The degree to which findings from in vitro-generated DCs reflect the in vivo-situation remains to be elucidated as well as the definite role of this consistent feature in the pathogenesis of JMML. Conclusion It seems conceivable that DCs derived from JMML MNC may contribute to clinical characteristics such as organ infiltration by ‘homing’ to DC-localizations/niches like the skin, spleen and liver. Acknowledgement This study was made possible by the support from the Zentrum für klinische Forschung I, University of Freiburg and from Investigating group "Molecular Biological System Transfer” (Iran). I would like to thank Prof. C. Niemeyer, Dr. U. Kontny and Dr. A. Mueller for the informative discussions and Prof. Dr. Ketelsen and Prof. Ig. Baumann for support of the electron microscopy studies. I would also like to thank our technicians Anja Lipowsky, Diana Birkle and Narges Amini for continuous support. References
© Copyright 2008 - TUMS PUBLICATIONS The following images related to this document are available:Photo images[pe08034t4.jpg] [pe08034t2.jpg] [pe08034f1.jpg] [pe08034t3.jpg] [pe08034t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}