 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
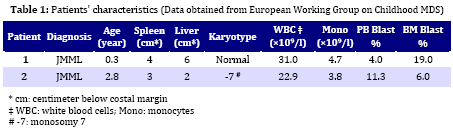
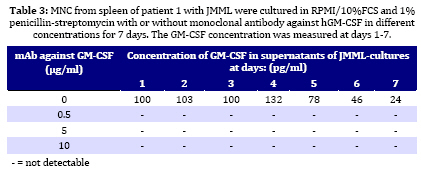
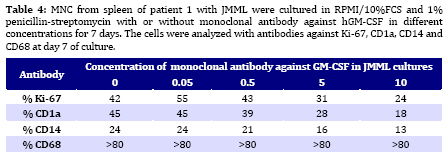
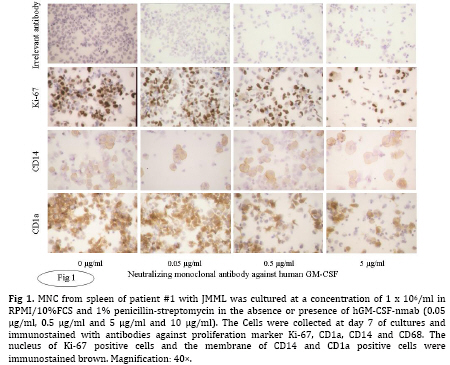
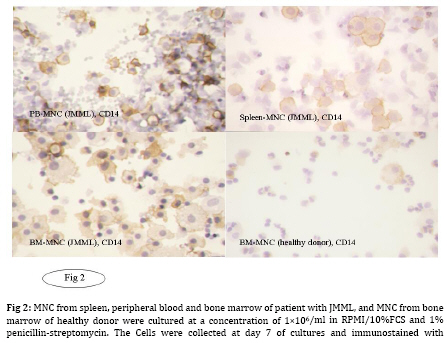
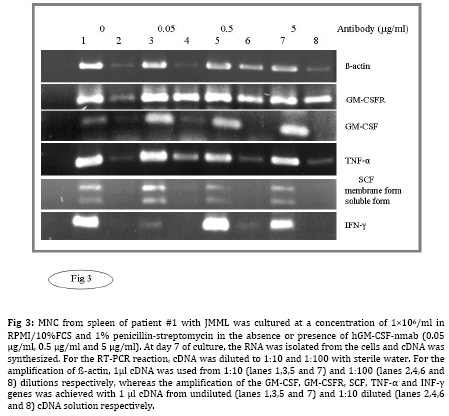
Iranian Journal of Pediatrics, Vol. 19, No. 3, Sept, 2009, pp. 213-233 The Role of GM-CSF in the Generation of CD1a Positive Cells in vitro by JMML Patients Parviz Shayan 1,2, DVM, PhD 1. Department of Pathobiology, University of Tehran, Tehran, IR Iran Received: Oct 11, 2008; Final Revision: Mar 29, 2009; Accepted: May 5, 2009 Code Number: pe09033 Abstract Objective: Juvenile myelomonocytic leukemia (JMML) is a rare myeloproliferative disorder of childhood. It was recently demonstrated that mononuclear cells from spleen, peripheral blood and bone marrow of the patients with JMML proliferate in vitro and population of the cells differentiate within 7 days to the CD1a positive dendritic cells. In the present study, the role of GM-CSF in the proliferation and differentiation of the JMML cells in vitro was investigated. Key Words: Juvenile myelomonocytic leukemia; CD1a; Dendritic cell; GM-CSF; Stem cell factor; Suspension cell culture; ELISA; Immunostaining; RT-PCR Introduction Juvenile myelomonocytic leukemia (JMML) is a rare myeloproliferative disorder in childhood. JMML is accompanied with leucocytosis and monocytosis, presence of elevated hematopoietic progenitor cells in the peripheral blood, massive infiltration of various organs causing skin rash, lymphadenopathy, hepatosplenomegaly and damage of lung tissue, which most probably lead to the death of patients[1,2]. The molecular mechanism involved in the pathogenesis of JMML is not well recognized. The mononuclear cells (MNC) isolated from spleen, peripheral blood (PB) or bone marrow (BM) of patients with JMML show spontaneous growth in vitro[3-6]. The cytokine responsible for this so-called “spontaneous growth” is reported to be GM-CSF. Emanuel et al 1991 have shown that the progenitor cells from JMML patients act hypersensitive to GM-CSF, which is produced by macrophages[3,7]. They showed that depletion of macrophages from the culture led to the loss of growth property of cells. Moreover, they have demonstrated that an antibody against GM-CSF also had an inhibitory effect in colony growth in the semi solid medium[7]. Beside GM-CSF, TNF-α seems to play also an important role in the proliferation of JMML cells. TNF-α neutralizing antibody is able to decrease the proliferation of the JMML in vitro semi solid culture[8, 9]. Furthermore, a tumor necrosis factor α specific ribozyme, which modulates GM-CSF gene expression, leads to decrease of GM-CSF-dependent colony formation and viability of JMML cells in vitro culture[10]. Analysis of GM-CSF receptor (GM-CSFR) showed that expression of GM-CSFR (count per cell) is not altered on the cells by JMML patients and nucleotide sequence analysis of the receptor also showed no novel mutation in the gene[11-13]. There is, however, evidence that proteins downstream of the GM-CSFR are involved in the pathogenesis of the disease[14]. Therefore, we previously analyzed the expression of hematopoietic cell phosphatase (HCP), a potential modulator of the GM-CSF-ras signal transduction pathway[15] and determined the ratio of intact interferon regulatory factor 1 and 2 (IRF-1, -2), which may contribute to the cell transformation[16] in patients with JMML. Our results showed that HCP transcript with or without exon skipping were not expressed differently in hematopoietic cells of patient with JMML compared to the healthy donors and neither tumor suppressor gene IRF-1 nor the oncogene IRF-2 was involved in the pathogenesis of JMML. Recently it was demonstrated that the MNC isolated from peripheral blood, bone marrow and spleen of the patients with JMML proliferate and differentiate to the CD1a positive cells in vitro in RPMI/10%FCS without exogenous growth factors[6]. In the present study, the role of GM-CSF in the proliferation and differentiation of the JMML cells in vitro was investigated. For this aim, the transcript of GM-CSF gene was first measured in the supernatant of the cell cultures using colorimetric Sandwich ELISA. Additionally, the GM-CSF was neutralized using monoclonal antibody against human GM-CSF and the effect of neutralization on the differentiation and proliferation of the JMML-cells was investigated in vitro. Furthermore, the expression of GM-CSF, GM-CSFR, TNF-α and SCF was determined in the antibody treated and untreated JMML-cells at mRNA level. Subjects and Methods Patients and controls: MNC from spleen of 2 patients with JMML were analyzed at the time of diagnosis. Clinical data of patients are shown in (Table 1). All tissues had been obtained with consent given according to institutional guidelines. Isolation of Mononuclear Cells: Mononuclear cells (MNC) were isolated by Ficoll- Paque (Pharmacia, Uppsala, Sweden) density gradient centrifugation. Spleen samples were cut into small pieces and placed in phosphate-buffered saline (PBS). The cells were isolated by scraping, filtering through a cell strainer (70 μm Nylon, Becton Dickinson) and by Ficoll-Paque. Cell culture: MNC from spleen were cultured at a concentration of 1x106/ml in RPMI medium with 10% FCS and 1% penicillin-strepto-mycin in culture dish. Cultures were incubated for 7 days at 37°C in atmosphere containing 5% CO2. At day 7 cells in suspension were counted and used for cytospin preparation. Immunocytochemistry: For immunostaining of cultured cells, we used an immunoperoxidase system from DAKO Hamburg, Germany. The used antibodies were CD1a (clone NA1/34), CD14 (clone TÜK4), CD68 (clone KP1) and Ki-67 (code No.A0047). These antibodies were purchased from DAKO, Germany. The number of positive reaction was determined for Ki-67, CD68, CD1a and CD14 in at least 1000 cells. Ki 67 was used as a proliferation marker. All proliferating cells expressed proliferating associated nuclear protein, Ki-67, whereas this protein was absent in the quiescent cells. Measurement of GM-CSF in supernatant of cultured MNC: MNC from spleen of 2 patients with JMML was cultured in RPMI/10%FCS for 7 days. For determination of GM-CSF, supernatant of cultures were centrifuged every second days at 17860 g and 4°C for 2 minutes, transferred to the new sterile tubes, aliquoted and stored at –20°C until used. GM-CSF was then measured using colorimetric Sandwich ELISA kits purchased from R&D Systems GmbH, Germany. Neutralization of GM-CSF in the cultures: The hGM-CSF neutralizing monoclonal antibody (hGM-CSF-nmab) was purchased from R&D Systems GmbH, Germany. This antibody has been selected for its ability to neutralize the biological activity of rhGM-CSF (R&D Systems). The ND50 for this lot of antibody was determined to be approximately 0.5-1.5 µg/ml in the presence of 0.5 ng/ml of rhGM-CSF using the factor-dependent human cell line TF-1 (R&D Systems). Since it was difficult to have enough MNC from PB or BM of the patients, the spleen was used as a source for the isolation of the MNC. MNC from spleen of patient #1 with JMML was cultured at a concentration of 1x106/ml in RPMI/10%FCS in the absence or presence of hGM-CSF-nmab (0.05 μg/ml, 0.5 μg/ml, 5 μg/ml and 10 µg/ml). The antibody was added to the culture once at the initiation time of culture. The cultures were judged daily under microscope. At day 7, MNC were collected and used for preparation of cytospin slides. The supernatants of the cultures were collected at days 1, 2, 3, 4, 5, 6 and 7 by centrifugation at 17860 g and 4°C for 2 minutes. Before measurement of GM-CSF concentration in the supernatant of the cell cultures, antigen (GM-CSF)-antibody complex was first removed from the supernatant. Briefly, 300 μl of the supernatant was incubated with Dynabeads M-450 coated with goat anti-mouse IgG (Dynal, Hamburg, Germany), which has been already washed with PBS, to remove the antigen-antibody-complex. After 1 h incubation on ice, the Dynabeads were removed using the Magnetic Particle Concentrator Dynal MPC-E (Dynal, Hamburg, Germany) and the GM-CSF concentration was determined as described above. Immunocytochemistry analysis of the cells was performed at day 7 of culture on cytospin as described above. Polymerase chain reaction (PCR): RNA from cultured cells was achieved according to the PegGold Trifast Kit (Peglab Biotechnologie GmbH, Erlangen Germany). One to 5 µg RNA was mixed with 2 pmol hexanucleotide (GibcoBRL/Life Techno-logies) in 12 µl DEPC. After a denaturation step at 70°C for 10 minutes, RNA was reverse transcribed into cDNA by incubating the 20 µl reaction containing 1 µl dNTP (10 mM each), 2 µl 0.1M DTT, 4 µl 5 x first strand buffer and 1 µl (200 U) superscript II (GibcoBRL/Life Technologies) at 40°C for 60 minutes. Enzyme activity was stopped by a final step at 70°C for 15 minutes. For RT-PCR reaction, cDNA was diluted to 1:10 and 1:100 with sterile water. For amplification of ß-actin, 1 µl cDNA was used from 1:10 and 1:100 dilution respectively, whereas amplification of other genes was achieved with 1 µl cDNA from undiluted and 1:10 diluted cDNA solution respectively. cDNA was amplified in a 100 µl volume containing 10 µl 10 x PCR-buffer, 3 µl 50 mM MgCl2, 2 µl dNTP (10 mM each), 2 µl sense primer (20 µM), 2 µl antisense primer (20 µM) and 0.5 µl (5000U/ml) Taq polymerase (GibcoBRL/Life Technologies). The nucleotide sequences of the different analyzed genes are listed as follow: β-actin sense 5` AAGAGATGGCCACGGCTGCT 3` antisense 5` TCGCTCCAACCGACTGCTGT 3` GM-CSF sense 5´ CTGCTGAGATGAATGAAACAG antisense 5´ GTGCTGCTTGTAGTGGCT 3´ GM-CSFR sense 5´ CCAGCAATGTCACCGTACGTT 3´ antisense 5´ TGTGGAACTGGCGGGAACAG 3´ SCF sense 5´ CCCAGGCTCTTTACTCCTGAAG 3´ antisense 5´ CTGCCCTTGTAAGACTTGGCTG 3´ TNF-α sense 5´ CAGAGGGAAGAGTTCCCCAG 3´ antisense 5´ CCTTGGTCTGGTAGGAGACG 3´ IFN-γ sense 5´ CAGGTCATTCAGATGTAGCG 3 antisense 5´ GCTTTTCGAAGTCATCTCG 3´ To avoid amplifying sequences of the ß actin pseudogene, the ß-actin primers were chosen from those DNA regions, which are absent in the pseudogene. The primer pairs were located on different exons so that the fragment length was indicative that exclusively cDNA was amplified. The amplification profile involved one pre-denaturation step at 95°C/3 minutes, denaturation at 95°C/ 45 seconds, annealing at 54°C/30 seconds, elongation at 72°C/45 seconds (35 cycles) followed by 10 minutes at 72°C after the last cycle (PCRExpress, Hybaid, UK). The PCR products were size fractionated by electrophoresis on 1-1.5% agarose gel in 0.5 x TBE (100 mM tris base, 100 mM boric acid and 2 mM EDTA pH 8.3), and stained with ethidium bromide. Findings Determination of GM-CSF in supernatant of JMML culture: MNC from two patients with JMML (pt #1 and pt #2) were cultured for 7 days in RPMI/10%FCS. One of these patients had monosomy 7 whereas the other had normal karyotype (Table 1). GM-CSF was measured every second days in supernatant of JMML-MNC cultured for 7 days. In the supernatants of both cultures, GM-CSF (pt #1 Ø 117 pg/ml; pt #2 Ø 56 pg/ml) could be determined (Table 2). The measured concentration of GM-CSF at day 1 in supernatant of in vitro culture of patient with normal karyotype was comparable to that of the patient with monosomy 7 (pt #1 76 pg/ml, pt #2 78 pg/ml). But at days 3, 5 and 7 of MNC-cultures, the concentration of GM-CSF in supernatant of the patient with monosomy 7 was lower than that of patient with normal karyotype (Table 2). Immunocytochemistry analysis of cultured cells from the patient 1 showed that 38% of the cells were Ki-67 positive. The percentage of CD1a and CD68 positive cells was 31% and more than 80% respectively. Immunocytochemistry analysis of cultured cells from the patient 2 showed that 24% of the cells was Ki-67 positive. The percentage of CD1a and CD68 was 8% and more than 80% respectively (data not shown). Neutralization of GM-CSF in JMML cultures: Neutralization of GM-CSF was performed in cultured MNC isolated from spleen of patient 1. GM-CSF was neutralized with 0.05 µg/ml, 0.5 µg/ml, 5 µg/ml and 10 µg/ml of monoclonal antibody against human GM-CSF. The amount of monoclonal antibody was calculated according to the concentration of GM-CSF in the cultures measured at days 1, 3, 5 and 7, and to the neutralizing capability of antibody demonstrated by R&D Company. After removal of GM-CSF-Ab-complex with beads-conjugated anti-mouse IgG, GM-CSF concentration was determined in supernatant of untreated as well as treated cultures with 0.5 µg/ml, 5 µg/ml and 10 µg/ml of monoclonal antibody against human GM-CSF at days 1 to 7. The concentration of GM-CSF in supernatant of untreated culture was 100 pg/ml, 103 pg/ml, 100 pg/ml, 132 pg/ml, 78 pg/ml, 46 pg/ml and 24 pg/ml for days 1 to 7, respectively (Table 3). Free GM-CSF could not be measured from day one to day seven in the supernatant of treated cultures with 0.5 µg/ml, 5 µg/ml and 10 µg/ml antibody. The count of the cells was in supernatant of the untreated culture at day 7, 1.15 x 106/ml, whereas it was decreased to 0.28 x 106/ml, 0.23 x 106/ml and 0.2 x 106/ml, when the cultures were treated with 0.5, 5 and 10 µg/ml antibody respectively. Immunocytochemical analysis of the cells at day 7 showed that the percentage of CD68 positive cells remained unchanged. The percentage of proliferating cells (Ki-67 positive) treated with 0.05 and 0.5 µg/ml antibody was comparable to that of untreated cultures (approximately 42%). The percentage of proliferating cells reduced to 31% or 24% in cultures treated with 5 µg/ml or 10 µg/ml antibody respectively (Table 4, Fig. 1). The percentage of CD1a positive cells remained unchanged in cultures treated with 0.05 µg/ml antibody compared to the untreated cultures. The CD1a positive cells decreased to 39%, 28% and 18%, when the cultures were treated with 0.5, 5 and 10 µg/ml antibody respectively. It was shown that MNC from normal healthy do not differentiate into the CD14 positive cells in abovementioned culture. Interestingly, we could observe that CD14 positive cells could be demonstrated at day 7 in previously some examined PB-MNC (>5%), BM-MNC (>20%) from patients with JMML (Fig. 2, data not shown). To determine the role of GM-CSF in the generation of CD14 positive cells by JMML patients,the phenotype of the cells in untreated and treated cultures was determined using monoclonal antibody against CD14 (Fig. 1). The percentage of CD14 positive cells remained unchanged (24%) in cultures treated with 0.05 µg/ml antibody compared to the untreated cultures, whereas it was decreased to 21%, 18% and 13% in cultures treated with 0.5, 5 and 10 µg/ml antibody respectively (Table 4). RT-PCR analysis was performed with the cells obtained from the untreated and treated cultures with 0.05, 0.5 and 5 µg/ml antibodies. The results showed that the mRNA for GM-CSF, GM-CSFR, SCF, TNF-α and INF-γexpressed in all examined cultures (Fig. 3). Discussion Molecular pathogenesis of JMML is rarely understood. It is known that JMML progenitor cells exhibit spontaneous colony formation in vitro. It is believed that selective hypersensitivity of myeloid progenitor cells to the GM-CSF plays a prominent role in the spontaneous growth of JMML granulocyte-macrophage colony forming units (CFU-GM) in semisolid culture[3,7] as well as in vivo[17]. A previous study[4-6] established a culture system of JMML MNC from PB, BM and spleen without exogenous growth factor supplementation. The cultures were characterized by marked adherence of cells and proliferative activity with frequent mitosis and a high Ki-67 index. Remarkably, after 5 days of culture, all JMML specimens featured differentiation into CD1a positive cells. Since the hematopoietic progenitor cells of JMML patients show a selective hypersensitivity to the GM-CSF, our observation allowed the speculation that the GM-CSF may be involved in the proliferation and differentiation of JMML cells into the CD1a positive cells in vitro. To investigate the role of GM-CSF in this culture system, the concentration of GM-CSF was determined in the supernatant of the JMML cultures. As expected, GM-CSF could be detected in the supernatant of both JMML cultures. The concentration of GM-CSF in the supernatant of the culture from JMML patient with monosomy 7 was lower than from the patient with normal karyotype. The importance of GM-CSF in the generation of CD1a positive cells can be inferred from the culture methods that allowed in vitro generation of DC from CD34+ precursors, usually by the sequential use of different cytokines combinations such as GM-CSF, IL-4 and/or TNF-alpha or SCF + IL-3 + IL-6 followed by GM-CSF + IL-4 [18]. Likewise, culture assays from monocytes have been described, using GM-CSF, IL-4 and TNF-α[19]. Since the importance of GM-CSF in the pathophysiology of JMML is unchallenged, the effect of GM-CSF in the proliferation and differentiation of JMML cells into population with DC like features was found out using different concentrations (0.05, 0.5, 5 and 10 µg/ml) of monoclonal antibody against human GM-CSF. To control the neutralizing effect of Antibody, GM-CSF concentration was measured every day in supernatant of the treated cultures. The monoclonal antibody at concentrations of 0.5, 5 and 10 µg/ml could neutralize the GM-CSF in all treated JMML cultures as measured by colorimetric Sandwich ELISA. In cultures treated with antibody concentrations of 0.5, 5 and 10 µg/ml, the total cell number decreased at day 7 at doses dependent manner. At the same time the percentage of CD68 positive cells remained unchanged, while the GM-CSF neutralizing antibody had decreasing effect at higher concentrations on the proliferation and generation of the cells into CD1a and CD14 positive cells. Although GM-CSF was not detectable in the cultures treated with monoclonal antibody, but its expression in the cells was demonstrable at the mRNA level. Therefore, it is to speculate that GM-CSF has been produced by macrophages and secreted into the treated culture constitutively. And very low amount of undetectable GM-CSF could be enough for the giving signal to the GM-CSF hypersensitive JMML progenitor cells to differentiate into the CD1a and CD14 positive cells. On the other hand, it was shown that on the cover slips CD1a positive cells were located in direct neighborhood and often with cell-to-cell contact with macrophages[6]. This observation led to speculate that the binding of the macrophages to the JMML haematopoietic progenitor cells may cause the macrophages to become polarized. The cytoskeleton is then reoriented to align the Golgi apparatus and the microtubule organizing center towards the JMML progenitor cells. GM-CSF secretion is thus specifically directed onto the progenitor cells, as it is the case by release of granules from the cytotoxic cells to the target cells. At the mRNA level the expression of GM-CSFR, TNF-α, SCF and IFN-γ was also demonstrable. Analysis of GM-CSF receptor (GM-CSFR) has shown that the expression of GM-CSFR (count per cell) was not altered on the cells by JMML patients and the nucleotide sequence analysis of the receptor showed also no novel mutation in the gene[11-13]. It was shown that TNF-α neutralizing antibody was able to decrease the proliferation of the JMML in semi solid medium[8,9]. Sawai et al 1996 reported that one possible mechanism for the cell proliferation in JMML is the strong proliferation of GM progenitors induced by haematopoietic factors, in particular SCF[20]. Conclusion TNF-α neutralizing antibody decreases also in the presented culture system the proliferation and differentiation of JMML progenitor cells into the CD1a positive cells (data not shown). It is obvious that the proliferation and differentiation of the cells to the CD1a positive cells in vitro liquid culture is poly factorial. The results in the present study suggest that GM-CSF plays an important role in the proliferation of the JMML cells in liquid culture. Furthermore, the results revealed that the GM-CSF is also important for the differentiation of the JMML progenitor cells into the CD1a and CD14 positive cells in vitro. Acknowledgment This study was made possible by the support from the Zentrum für klinische Forschung I, University of Freiburg, Müller-Fahnenberg-Stiftung and Investigating group "Molecular Biological System Transfer" (MBST, IRAN). I would like to thank Prof. Dr. Niemeyer at Division of Pediatric Hematology and Oncology, Department of Pediatrics and Adolescent Medicine, University of Freiburg, Mathildenstrasse, Germany, for the informative discussions, Prof. Dr. Martin at Medizinische Hochschule Hannover, Pharmakologie OE 5322, and Dr. HH. Seydewitz at the University Children's Hospital, University of Freiburg for the measurement of GM-CSF in supernatant of cultures. References
© 2009 by Center of Excellence for Pediatrics, Children’s Medical Center, Tehran University of Medical Sciences, All rights reserved. The following images related to this document are available:Photo images[pe09033f3.jpg] [pe09033f1.jpg] [pe09033t2.jpg] [pe09033t3.jpg] [pe09033t1.jpg] [pe09033f2.jpg] [pe09033t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}