 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
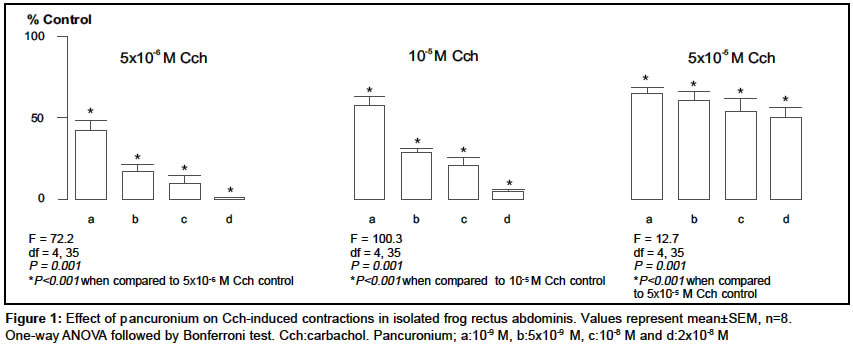
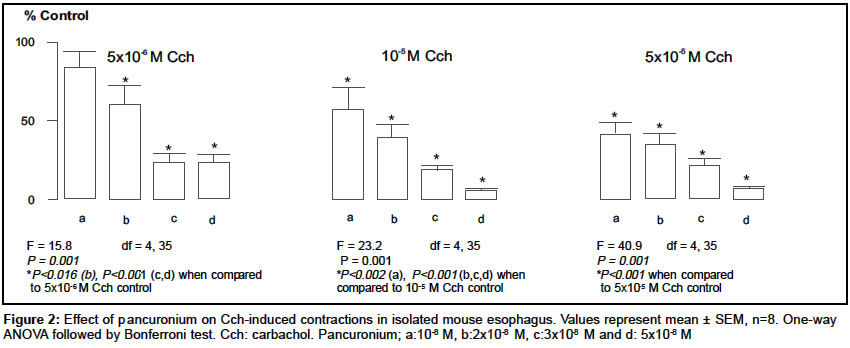
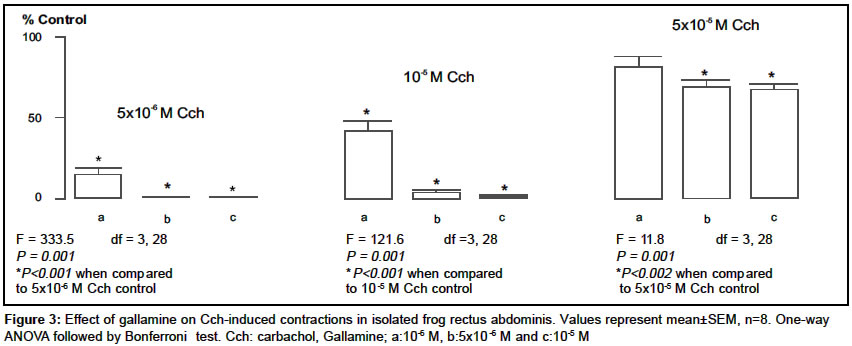
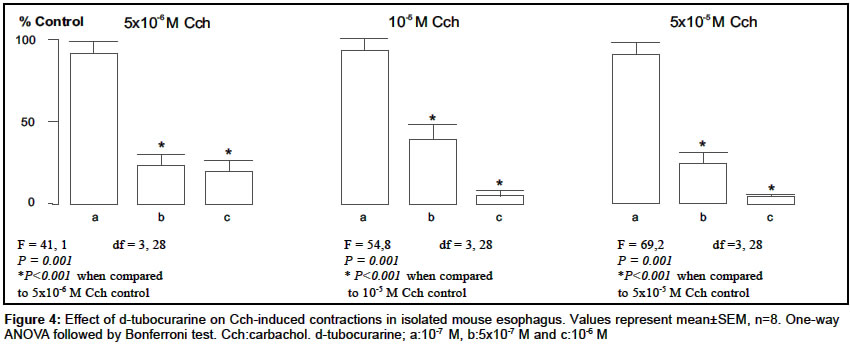
Indian Journal of Pharmacology, Vol. 36, No. 2, April, 2004, pp. 72-75 Research Paper Characterization of nicotinic cholinergic receptors Döndas Naciye , Karatas Y, Dikmen A Department of Pharmacology, Medical Faculty, Çukurova University, 01330 Adana Code Number: ph04028 ABSTRACT Objective: To characterize nicotinic cholinergic receptors (cholinoceptors) in frog rectus abdominis and mouse esophagus. Material and Methods: Isolated preparations of mouse esophagus and frog rectus abdominis were separately mounted in organ baths filled with Krebs and Ringer solutions respectively. The organ bath medium was continuously gassed with 95% O2 and 5% CO2 and the temperature was maintained at 37°C and 25°C for mouse esophagus and frog rectus abdominis respectively. Changes in muscle length were recorded by isotonic transducers. The effects of neuromuscular blocking agents were investigated on contractions mediated by nicotinic cholinoceptors in both the preparations. Results: The contractions induced by carbachol (Cch) (5x10-6-5x10-5 M) were significantly inhibited by pancuronium (10-9-5x10-8) in mouse esophagus and frog rectus abdominis. Gallamine inhibited the contractions in the frog rectus abdominis but it failed to reduce the Cch responses in the mouse esophagus. On the other hand, d-tubocurarine caused irreversible contractions in the basal tonus of frog rectus abdominis, but it reduced the contractions evoked by Cch in mouse esophagus. Conclusion: The different effects of the neuromuscular blocking agents on contractions mediated by nicotinic cholinoceptors in frog rectus abdominis and mouse esophagus indicated that the nicotinic cholinoceptors in two different tissues may not be identical and these differences may be attributed to the different nicotinic cholinoceptor subtypes.INTRODUCTION In the periphery, nicotinic cholinoceptors are distributed at neuromuscular junctions and ganglionic synapses. The molecular structure of these receptors is very similar but they differ pharmacologically,[1] which is attributable to the existence of several variants of a-,b-, and g-subunits.[2] Curariform drugs can block neuromuscular transmission by interfering with the postsynaptic action of nicotinic cholinoceptors (NM) at the skeletal muscle. These drugs have a different mode of action[3] depending upon the molecular properties of the drug and inherent properties of the myoneural junction. Pancuronium, gallamine and d-tubocurarine are curarimimetic neuromuscular blocking agents and act as competitive antagonists at nicotinic acetylcholine receptors of the motor endplate. Nicotinic cholinoceptors have been identified in frog rectus abdominis[4] and mouse esophagus.[5] It was reported that d-tubocurarine[6] and gallamine[7],[8] interact with these receptors at the skeletal muscle. Pancuronium caused neuromuscular depression in mouse phrenic nerve-diaphragm preparation.[9] It was also suggested there are some differences between nicotinic cholinoceptors in different species of animals.[10] The present study was conducted with an aim to evaluate the characterization of nicotinic cholinergic receptors in frog rectus abdominis and mouse esophagus. MATERIAL AND METHODS AnimalsAnimals used in the present study were obtained from the Çukurova University Medical Sciences Research Centre, Adana, Turkey. Both sexes of mice (Mus musculus var. Albino) weighing 20-30 g were housed in polypropylene cages in controlled temperature (27 ± 2°C) and light. They were fed with standard laboratory pellets. Food and water were provided ad libitum. Frogs (Rana pipiens) weighing 15-25 g were housed in pools in a regulated environment. The principles of laboratory animal care published by NIH were followed during the experiments and the study was approved by the Committee of Ethics at the Çukurova University Medical Sciences Research Centre. Drugs and chemicals All the chemicals were dissolved in distilled water and made up to a volume of 0.1 ml to instill in the organ bath. Tissue preparations b) Frog rectus abdominis: The rectus abdominis muscle was removed after the animal was pithed and decapitated. The abdominal skeletal muscle was divided into two halves by dissecting along the linea alba. The two preparations were separately mounted under 0.5 g tension in jacketed organ baths containing Ringer solution [(as mM: NaCl 111.1, KCl 1.87, CaCl2 1.08, NaH2PO4.2H2O 0.08, NaHCO3 2.38, Glucose 11.1)]. The temperature of the bathing medium was maintained at 25°C and the medium was continuously aerated with 95% O2 and 5% CO2. The tissue was equilibrated for 1h before starting the experiment. Changes in the muscle length were recorded on a smoked drum via an isotonic lever (x8-10 magnification). Experimental protocols Statistical analysis RESULTS Carbachol (Cch; 5x10-6, 10-5 and 5x10-5 M), the mixed muscarinic-nicotinic acetylcholine receptor agonist, produced concentration-dependent contractions (in mm) in both tissues (n=10) and there was no significant difference between the first and second series of the control group in mouse esophagus (first series, 3.8 ± 0.6, 10.4 ± 1.3 and 19.9 ± 1.5; second series, 4.6 ± 0.9, 10.9 ± 2.0 and 21.1 ± 2.3 respectively) and frog rectus abdominis (first series, 7.5 ± 0.7, 24.0 ± 1.9 and 55.7 ± 3.6; second series, 7.6 ± 1.6, 25.9 ± 2.1 and 60.2 ± 3.8 respectively). Pancuronium (10-9-5x10-8 M) significantly reduced Cch contractions in both the tissues. However, the nature of this inhibition was not identical; the inhibitory effect of this agent was partially reversed by the higher Cch concentrations (10-5 and 5x10-5 M) in frog rectus abdominis [Figure - 1] but not in mouse esophagus [Figure - 2] . Gallamine (10-6, 5x10-6 10-5 M) inhibited the contractions induced by Cch in the frog rectus abdominis [Figure - 3] , but it had no effect on Cch-responses in the mouse esophagus in the absence (as % control: 102.2 ± 6.3, 94.6 ± 9.9 and 95.3 ± 7.1 respectively for each Cch concentration) and presence (as % control: 104.3 ± 5.8, 99.8 ± 6.3 and 102.8 ± 5.9 respectively for each Cch concentration) of atropine (10-6 M). d-tubocurarine (10-7, 5x10-7, 10-6 M) significantly reduced the Cch-contractions in the mouse esophagus in a concentration-dependent manner [Figure - 4] . On the other hand, it caused irreversible contraction in the basal tonus of the frog rectus abdominis muscle. Thus it could not be examined in this tissue. DISCUSSION In the present study, we compared the nicotinic cholinoceptors in mouse esophagus and frog rectus abdominis. The intact mouse esophagus includes nicotinic and muscarinic cholinoceptors. And muscarinic cholinoceptors are confined to the distal 0.5 cm of the tunica muscularis externa.[11] To obviate the muscarinic cholinoceptors in mouse esophagus we used the mucosal tissue-free, upper 2/3 of esophagus and this was checked with atropine, a selective antagonist of muscarinic cholinoceptor which has no effect on Cch-induced contractions. Pancuronium (10-9-5x10-8 M) significantly inhibited the Cch-induced contractions in the mouse esophagus and frog rectus abdominis in a concentration-dependent manner [Figure - 1] and [Figure - 2] . However, the nature of this inhibition was not identical in the two different tissues. For example, in the frog rectus abdominis, the inhibitory effect of pancuronium was partially reversed by higher Cch concentrations (10-5 and 5x10-5 M), which is consistent with competitive inhibition by this drug. But in the mouse esophagus pancuronium progressively inhibited the Cch-contractions and this effect was not reversed by higher Cch concentrations (10-5 and 5x10-5 M). Similar results were observed with gallamine in the frog rectus abdominis [Figure - 3] . However, gallamine failed to inhibit the Cch-responses in the mouse esophagus, possibly due to lack of penetration of gallamine into the mouse esophageal tissue since these drugs are all quaternary ammonium compounds. On the other hand, d-tubocurarine caused irreversible contraction at the basal tonus of the frog abdominal skeletal muscle. The cause of this effect may be due to the non-specific action of it on this tissue. However, d-tubocurarine significantly inhibited the Cch-responses in the mouse esophagus [Figure - 4] . The nature of the action of this drug was similar to that of pancuronium in the same tissue. It did not act as a competitive neuromuscular blocker in the mouse esophagus. Considering these observations, the question arises as whether these neuromuscular blocking agents are competitive inhibitors only in the frog rectus abdominis but not in the mouse esophagus. It was reported by Ramaswamy et al (1978) that Cch releases acetylcholine (ACh) from the presynaptic nerve terminals by depolarizing the presynaptic nicotinic receptors in the skeletal muscle.[12] A similar state might have occurred in our study and thus Cch may stimulate ACh release from the intramural nerve plexus. So Cch-induced ACh release may also have a role in the interaction between Cch and neuromuscular blocking agents towards nicotinic cholinoceptors. In the frog rectus abdominis this contribution may be stronger than in the mouse esophagus. The different effects of neuromuscular blocking agents in two different tissues are likely to be due to a species difference, or different subtypes of nicotinic cholinoceptors. This is in agreement with the report of Bonta et al (1968), who suggested that pancuronium was more potent in producing blockade on the tibialis, a white muscle, than on the soleus, a red muscle and this agent is different in this respect from the prototype competitive blocker d-tubocurarine.[13] Additionally, Kerr et al (1995) reported the differences in nicotinic cholinoceptor subtypes involved in vagal ganglionic responses.[10] These experimental data are consistent with our results and support our suggestion about different nicotinic cholinoceptor subunits in the mouse esophagus and frog rectus abdominis. In conclusion, the results reveal that the effects of neuromuscular blocking agents on contractions mediated by nicotinic cholinoceptors were different in the mouse esophagus and frog rectus abdominis. These differences may reflect the different nicotinic cholinoceptor subtypes in the two tissues. ACKNOWLEDGEMENTS We sincerely acknowledge Çukurova University Research Foundation for providing the financial assistance to conduct the study and we also would like to thank Edward Neale, Tim Munsey and Dr. Malcolm Hunter for their valuable suggestions.REFERENCES
Copyright 2004 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph04028f4.jpg] [ph04028f2.jpg] [ph04028f3.jpg] [ph04028f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}