 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
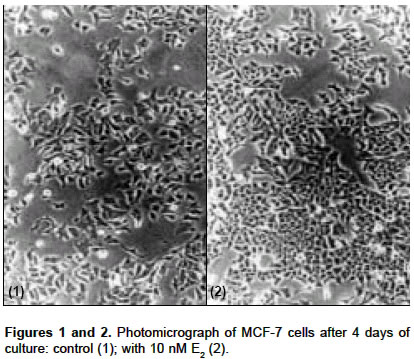
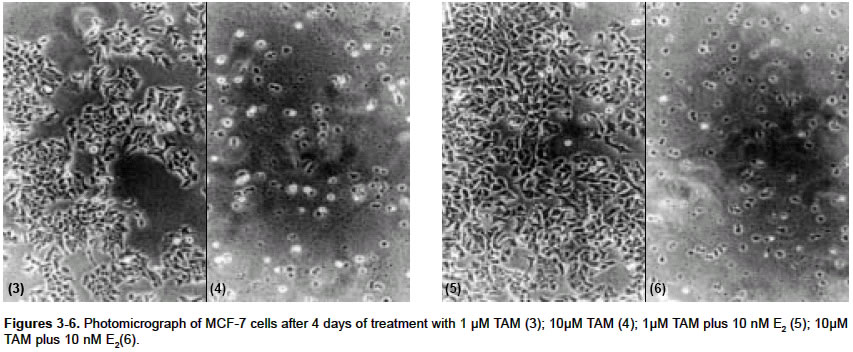
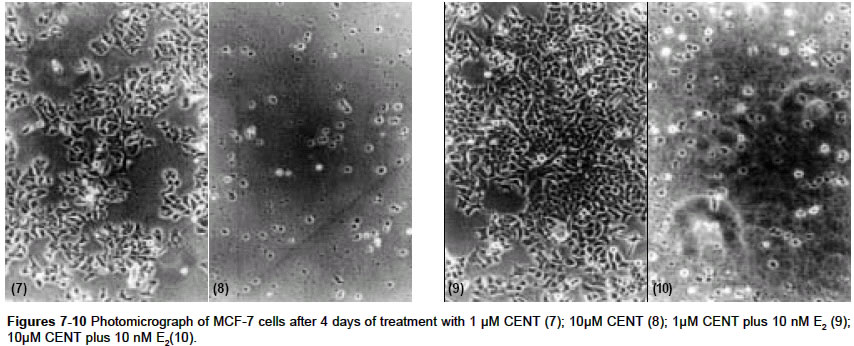
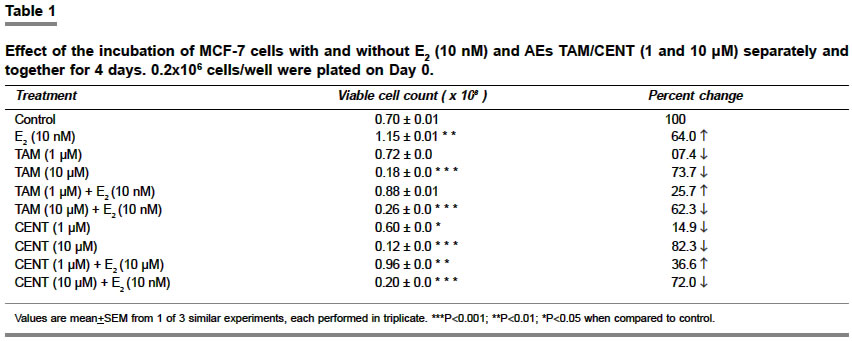
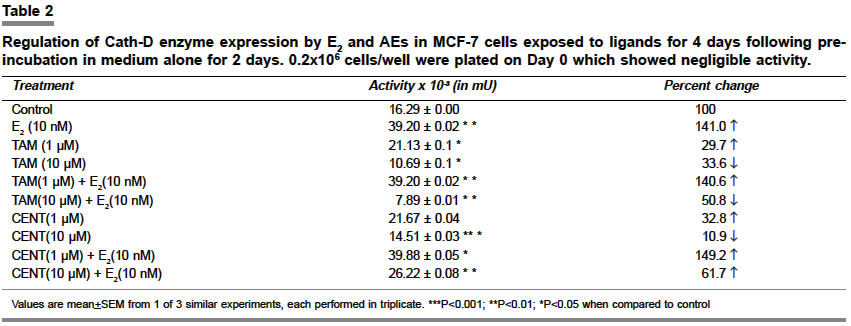
Indian Journal of Pharmacology, Vol. 36, No. 4, August, 2004, pp. 238-243 Research Paper Morphological and biochemical basis of centchroman as a novel antineoplastic agent in MCF-7 human breast cancer cells Srivastava S, Sharma R, Balapure Anil K Tissue Culture Laboratory, National Laboratory Animal Centre, Central Drug Research Institute, Lucknow - 226 001 Code Number: ph04078 ABSTRACT OBJECTIVE: To investigate the antineoplastic potential of a novel antiestrogen (AE) centchroman (CENT) using cell morphology, viability and cathepsin-D (Cath-D) gene expression as indices in MCF-7 human breast cancer cell line. MATERIAL AND METHODS: MCF-7 cells were incubated with CENT (1 µM, 10 µM) in the absence and presence of 17- estradiol (E2, 10 nM) for 4 days. Investigations were carried out on alterations in cell morphology, number, Cath-D gene expression using Phase Contrast Microscopy, Trypan-blue dye exclusion and spectrophotometric assay. A standard antiestrogen and antibreast cancer agent tamoxifen (TAM -1 µM, 10 µM) was used as a positive control. RESULTS: E2 (10 nM) alone significantly up-regulated the cells morphologically, numerically and biochemically thus establishing their hormonal responsiveness. TAM and CENT at 1 µM each enhanced and reduced the viable cell number respectively whereas both elevated the Cath-D activity. However, 35mm photographic evidence did not indicate any cytotoxic effect with either drug as compared to control. Addition of E2 to the two ligands separately affected the three parameters similar to that in E2 alone. With 10 µM TAM and CENT individually, the cells were visually devastated and so were the viable cell numbers and Cath-D gene expression. Supplementation of E2 under similar conditions failed to undo the damage to the cells morphologically and quantitatively. However, disparate results were observed with regards to the enzyme activity where TAM+E2 caused inhibition and CENT+E2 resulted in an induction. CONCLUSION: It can be concluded that both the AEs TAM and CENT displayed similarities in their antineoplastic action on MCF-7 cells as observed by morphological and viability studies except that the latter proved cytotoxic even at 1µM concentration. TAM and CENT showed a similar effect on Cath-D gene expression except at the higher dose (10 µM). These results suggest that the antiestrogen CENT has better antineoplastic potential as opposed to TAM. Keywords: 17-b estradiol, centchroman, tamoxifen, neoplastic INTRODUCTION Estrogen (E2) initiates and regulates a cascade of events following interaction with the estrogen receptors (ERs) which are copiously expressed in human breast cancer-derived cell lines such as MCF-7, T47D and ZR75.[1] The MCF-7 cell line, a human breast adenocarcinoma consisting of epithelial cells isolated from the pleural effusion of a post-menopausal female, has thus far provided an ideal tool for us[2] and others[3],[4] for evaluating E2-dependent phenomena in vitro. This has significantly led to the identification of tamoxifen (TAM) and other antiestrogens (AEs) for breast cancer therapy.[5],[6],[7] Besides, other estrogen-related phenomena such as the expression of proteases and antiproteases which prima facie regulate the invasion and metastasis, have also been investigated using MCF-7 and other ER-positive cell lines. One such enzyme cathepsin-D (Cath-D), an acidic lysosomal aspartyl endoprotease ubiquitously present in all the cells, constitutively expressed in ER-negative cells, and over-expressed in ER-positive breast cancer cell lines,[8] has attracted enormous research attention. In MCF-7 cells, Cath-D mRNA, tightly regulated transcriptionally by E2 and indirectly by growth factors[9] has a generally proven role in the invasion and metastasis of breast cancer. In ER-positive cell lines, the pro-Cathepsin-D mRNA is over-expressed, resulting in the accumulation of the active protease in large endosomes and the secretion of the precursor (pro-Cath-D, 52KD). In the lysosomes, the precursor is cleaved to the 48KD intermediate enzyme which is activated into two-chain mature form containing subunits of 34KD and 14KD respectively. However, in ER-negative breast cancer cells like MDA-MB 231, HMT 3522, T4-2, and HBL-100 Cath-D is constitutively expressed.[9],[10] A number of AEs, viz. TAM; ICI 182,780; raloxifene and EM-652 have been evaluated with respect to Cath-D vis-à-vis MCF-7 cells.[11] In the present study, we have investigated the morphological and numerical changes in MCF-7 cells vis-à-vis E2, Tamoxifen and a novel AE Centchroman (CENT).[11] Besides, the same ligands have been employed to assess the gene expression of Cath-D and its regulation as a biochemical marker. TAM is a well-known AE which has been used as a positive control. CENT, developed by this institute and hitherto found similar in its antiestrogenic properties to TAM, has been investigated for its antiestrogenic and antineoplastic potential. MATERIAL AND METHOD Material: 17-b estradiol (E2), TAM, hemoglobin, Bovine Serum Albumin (BSA), Dulbecco′s Modified Eagle′s Medium (DMEM), N-[2-Hydroxyethyl] piperazine-N′-2-ethanesulfonic acid (HEPES), penicillin, streptomycin, gentamicin and disposables were purchased from Sigma Chemical Co. St. Louis, MO, USA. Fetal Calf Serum (FCS) was purchased from GIBCO BRL Laboratories, New York, U.S.A. Cell culture: MCF-7 cells obtained from the National Centre for Cell Science, Pune, India and being maintained in our laboratory were cultured as reported previously by us.[2] Briefly, DMEM pH 7.4, containing penicillin (100 U/ml), streptomycin (100 µg/ml) and gentamicin (60 µg/ml) supplemented with 10% FCS and 10 mM HEPES was used for culturing the cells. This medium was labeled Medium I. The cells were cultured and subcultured in T-25 tissue culture flasks at appropriate intervals in a humidified atmosphere of 5% CO2 at 37°C in a CO2 incubator. For the purpose of experiments, the cells were cultured for a total of 6 days in a six-welled plate (35 mm diameter/well). For the initial 2 days, phenol red-free DMEM abbreviated as Medium II containing 10% FCS, stripped with Dextran Coated Charcoal was used (DCC/FCS). This was done to deplete the cells of steroids including E2 present in the serum and was prepared essentially according to Soto and Sonnenschein (1985).[12] This medium was utilized to evaluate the effect of extraneously added ligands such as E2, TAM and CENT for 4 days. Estrogen, antiestrogenic ligands treatment and viability studies: At the end of the incubation, each well was trypsinated and the trypan-blue dye exclusion technique performed to determine the ratio of live versus dead cells as reported previously.[2],[14] Cathepsin-D assay: The trypsinated cells were pelleted at 100 x g for 10 min at 4°C and re-suspended in 1 ml of chilled 1.15% KCl. The cells were lysed on ice in a Virtis sonicator with five short bursts of 10 seconds each. The quantitative expression of Cath-D activity in the lysate was conducted essentially according to Barret and Heath (1977)[15] employing hemoglobin as a substrate. Protein was quantified in the lysate as per the procedure of Lowry et al[16] using BSA as standard. One unit of the enzyme activity was defined as µMoles/min of the product generated from the substrate. Student′s ′t ′ test was used to determine the level of significance and a P value <0.05 was regarded as significant. RESULTS Morphological studies: The cells were examined daily under a Nikon Diaphot Phase-Contrast Microscope and photographed on Day 6 which marked the culmination of the experiment. The details of their morphological characteristics upon treatment with different ligands and comparison with controls are as follows. [Figure - 1] depicts the control (without any treatment) cells on Day 6 with a typical oblong, polygonal appearance glued to the substratum in a flattened cobblestone fashion. Abundant cytoplasm was observed in the entire cell populace which contained nuclei. Interspersed between the cells were occasional gap junctions signaling that the cells were in the replicative mode. The circumstantial presence of rounded dead cells was also noticed, floating in the medium in the background, which were removable upon replacement of the medium with fresh medium. Addition of 10 nM E2 [Figure - 2] displayed massive proliferative activity in a matter of 4 days confirming the steroid dependency of the cell line. Supplementation of the medium with TAM (1 µM, 10 µM), a well-known AE employed as a positive control in this study, resulted in a plethora of changes in the overall cellular architecture with more pronounced changes at the latter dosage. At 1 µM [Figure - 3] the cells seemed reduced both in size and density as compared to the control [Figure - 1]. With 10 µM TAM [Figure - 4], dramatic changes were observed where the cell population showed residual shrunken entities. The potential of 10nM E2 to synergise with or antagonize TAM at the two concentrations was also examined. 1 µM TAM plus 10 nM E2 [Figure - 5] displayed not only the complete reversal of the deleterious effect of the drug but a restoration of cell density similar to that observed with E2 alone as in [Figure - 2]. On the contrary, 10 nM E2 along with 10 µM TAM [Figure - 6] provided results akin to those observed with 10 µM TAM alone [Figure - 4]. The test AE CENT at 1 µM [Figure - 7] provided results similar to that of control [Figure - 1] and TAM at identical molarity [Figure - 3]. However, a near complete destruction of cells was noticed at 10 µM concentration [Figure - 8] presenting a scenario similar to that of TAM at identical concentration [Figure - 4]. Addition of 10 nM E2 in the presence of 1 µM CENT [Figure - 9] demonstrated the cellular profile similar to that observed with E2 alone [Figure - 2]. Subsequently, addition of 10 nM E2 along with 10 µM CENT [Figure - 10] failed to rectify the detrimental effect of the drug similar to that noticed with CENT or TAM alone as in [Figure - 8] and [Figure - 6] respectively. The observations thus far are clearly indicative of the antiestrogenic and antineoplastic potential of the drug. Cell viability: The purpose of this study was to ascertain the proliferative potential of cells in the absence and presence of E2 and its regulation by the AEs such as TAM and CENT. The results of these studies are collated in [Table - 1]. The increase in cell numbers from 0.2x106 (Day 0) at the time of plating to 0.7x106 on Day 6 (100%) under the conditions of incubation (2+4 days) confirms the normal proliferative potential of the cell line. Additionally, 10 nM E2 significantly upregulated the cell number by 64% over Day 6 control, demonstrating the estrogen responsiveness of the cell line in agreement with others.[17], [18] TAM displayed heterogeneous dose-dependency of MCF-7 cells′ responsiveness at 1 and 10 µM. Albeit a small and statistically insignificant increase in the cell number was noticed at 1 µM, a ten-fold higher concentration i.e. 10 µM caused a near complete devastation of the cell population (73.7% of control). One µM TAM was partially able to check the proliferative potential of 10 nM E2 when added together. Conversely, TAM at 10 µM in the presence of E2 precipitously declined the cell number to approximately 62% of control establishing its role as an AE. One µM CENT proved more effective than TAM at a similar concentration, significantly depleting the cell number by about 14%. Supplementation with 10 nM E2 at this dose countered the cytotoxicity of CENT and a modest but significant increase in cell number by 36.5% was observed. At higher concentration, CENT (10 µM) precipitously decreased the cell number by 82.2% which was not favorably reversible upon the addition of E2 (72% of control). Cathepsin-D activity: The ability of E2 and AEs to modulate the gene expression of the enzyme in terms of its activity was examined under the incubation conditions discussed beforehand and is summarized in [Table - 2]. It was observed that 10 nM E2 catapulted the enzyme activity from 16.29 mU to 39.20 mU, a massive (P<0.01) increase of 141%. TAM at 1 µM raised the induction to 29.7% as compared to control which was completely abolished at 10 µM dose where an overall decline of 33.6% was observed. The ability of 10 nM E2 to attenuate or synergise with the enzyme activity in the presence of TAM was also examined. At 1 µM, TAM was unable to inhibit the induction of activity where an increase of about 140% was recorded as compared to the control. It is, however, interesting to note that the percentage of induction by a combination of TAM and E2 or E2 alone was of the same order i.e. 141% as compared to the control taken as 100%. In other words, TAM at 1 µM failed to exert any antiestrogenic or estrogenic effect in the presence of E2. Subsequently, at 10 µM, TAM under similar conditions lowered the activity by 50% as compared to control. The test AE CENT at 1 and 10 µM behaved similar to TAM enhancing and diminishing the enzyme activity by 32% and 10% respectively. However, the extent of inhibition with CENT was significantly lesser as compared with TAM (33.6%) at 10 µM, possibly hinting at their dissimilar potential in regulating the enzyme gene expression. CENT at 1 µM and 10 µM in the presence of E2 provided further evidence of heterogeneity of actions, increasing the enzyme activity by 149% and 62% respectively as compared to control unlike TAM. DISCUSSION The surging incidence of breast cancer nationally and internationally has spurred the efforts to identify its prognostic and diagnostic markers as well as to improve the therapeutics available for its management. Cathepsin-D (Cath-D), a 34KD protein, is one such lysosomal marker protease naturally overexpressed in both estrogen receptor (ER)-positive and negative breast tumors.[8] Cath-D augments cell growth and decreases contact inhibition rather than cell invasion through the basement membrane, thereby promoting clinical metastasis.[8],[19] Cath-D is expressed by ER-positive, estrogen-dependent MCF-7 human metastatic breast cancer cell line and is estrogen-inducible. This study examined the inducibility of Cath-D gene expression by estrogen and its regulation by antiestrogens (AEs) such as TAM and CENT. In addition, morphological and cell viability studies conducted with phase-contrast microscope under similar conditions were also undertaken. To the authors′ knowledge, this is the first such report where the Cath-D expression has been examined vis-à-vis MCF-7 cells and CENT. The morphological and cell viability studies clearly indicate the estrogenic dependence of MCF-7 cells and the reversibility of the hormonal action by the test AE CENT. TAM, a standard AE has been used as a positive control by us and others.[5],[6],[20] Comparison between untreated Day 0 control cells (0.2x106 plated) versus Day 6 cells [Figure - 1] microscopically (100 x magnification) indicated an increase in the cell population consistent with the results of the viability studies [Table - 1]. The gross morphology of the cells indicated classical architecture with a well-defined plasma membrane, nucleus, and nucleoli in each cell. Estrogen at 10 nM established the hormonal responsiveness of the cells when the cell number rose up significantly by 64% [Table - 1] in 4 days, a fact attested by pictorial proof [Figure - 2]. TAM at 10 µM [Figure - 4] unequivocally demonstrated its antiestrogenic action rather than at 1 µM [Figure - 3], at which concentration a small, albeit statistically non-significant increase was observed. The lack of cent percent antiestrogenicity of TAM and similar compounds at low molarities such as 1 µM in this case is due to their belonging to Type I class of mixed AEs[11], [21],[22],[23] which are known to exhibit an estrogenic effect at low doses. CENT, which is a molecule belonging to an identical class, however, showed a slight disparate effect by lowering the cell number by approximately 15% at the lower dosage [Table - 1], [Figure - 7]. This hints towards the better efficacy of the compound in terms of restricting the cell growth even at low doses. At a tenfold higher dosage i.e. 10 µM [Figure - 8] its action was found to be similar to TAM. This, therefore, suggests that CENT besides possessing contraceptive activity[24], [25] has a significant amount of antineoplastic effect on human breast cells at both low and high doses. The examination of the ability of 10nM E2 to synergise with or antagonize the two AEs revealed interesting results. Both TAM and CENT at 1 µM each harmonized with the hormone E2 in augmenting the cell number with a significant rise in the latter case [Table - 1]. Clarke et al [26] have recently reviewed in detail the estrogenic nature of TAM at a low dose which may also hold true for CENT. The higher estrogenic effect of CENT rather than TAM at 1 µM in the presence of E2 resulting in increased mitogenesis implies disparity between the two ligands. A perusal of [Figure - 5] and [Figure - 9] seemingly confirms these observations. At the next higher molarity tested i.e. 10 µM, the two AEs exerted significant cytolytic activity in the presence of 10 nM E2. These observations are also corroborated by the pictures [Figure - 4] and [Figure - 8] and support the AE activity of these compounds. Perry et al (1995)[27] have also demonstrated the weak estrogenic nature of TAM at low dose (1 µM) and the cytocidal effect at 10 µM which support our observations. From the foregoing, it can also be derived that CENT has a higher degree of estrogenicity at 1 µM whereas it behaves similar to TAM at a 10 µM dose, thereby indicating the dissimilarity in their modes of action. The morphological evidence established with the viability data expectedly correlated with the biochemical parameter of Cath-D activity and its induction with E2. The gene for Cath-D is constitutively expressed by ER-negative cells such as MDA-MB 231 cell line and is induced by E2 in ER-positive cell lines such as MCF-7, T47D and ZR75.[8] [Table - 2] shows basal gene expression of Cath-D, its selective inducibility with E2, and 1 µM TAM, and complete abrogation at 10 µM-observations supported by several studies.[1],[28] E2 at 10 nM administered along with 1 µM TAM propped up the induction to the same extent as E2 alone whereas complete inhibition was observed at 10 µM concentration for reasons explained earlier. The lack of the additive effect on enzyme induction between 1 µM TAM and 10 nM E2 added together versus E2 alone needs to be probed further. A plausible explanation could be that the hormone achieves the maximal induction of the Cath-D gene leaving no room for TAM to exert its estrogenic effect. CENT behaved partially similar to TAM vis-à-vis Cath-D activity when evaluated alone at both the concentrations. Similarly, in the presence of E2, CENT at a lower concentration behaved like TAM whereas at a higher concentration the effect was opposite. This anomalous response of E2+10 uM CENT vis-à-vis TAM at similar dose may be specific only for Cath-D expression since morphological observations [Figure - 6] and [Figure - 10] and cell number values [Table - 1] show parity. A latest report suggesting Cath-D augmenting apoptosis in T lymphocytes[29] which is an unlikely property of the enzyme makes it imperative to conduct more studies to dissect out the dissociation between these parameters in relation to CENT. Similar cases of non-conformity between antiestrogenic and antigrowth factor properties of AEs have earlier been documented.[23] AEs such as ICI 164,384 and RU 486 are known to modulate Epidermal Growth Factor (EGF), Insulin like Growth Factor-I (IGF-I) etc. These do not necessarily require the presence of ER. Perry et al [27] have also reportedly determined that TAM exerts its antiestrogenic effect through ER and non-ER routes. A similar situation may also prevail with CENT which needs to be explored. In conclusion, our studies clearly demonstrate the efficacy of CENT as an AE which displays dose-dependent effects similar to TAM. The morphological evidence of CENT′s action on MCF-7 cells bears a complete resemblance with TAM whereas the data on cell viability has some exceptions. Absolute agreement was also noticed in the expression of the biochemical marker except at the 10 µM dose in presence of E2 where the former induces rather than inhibits Cath-D gene expression. A recent report on AE action[24] which has thoroughly reviewed the progress in this area has concluded that AEs with and without ER involvement may alter intracellular proteins, signaling pathways, oxidative stress response, protein kinase C, isoform activation, calmodulin function and plasma membrane architecture and function to exert their effect. A detailed examination on deciphering the molecular basis of CENT action is underway. This may identify the true antineoplastic potential of this novel entity despite TAM being the most widely used molecule in this class.[19] ACKNOWLEDGEMENT The authors would like to thank Dr C.M. Gupta, Director, CDRI for providing Research Assistantship to Swasti Srivastava and for displaying avid interest in this work. They would also like to acknowledge Dr S. Ray for providing the Centchroman sample. REFERENCES
Copyright 2004 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph04078f1.jpg] [ph04078f1-2.jpg] [ph04078t2.jpg] [ph04078f7-10.jpg] [ph04078f3-6.jpg] [ph04078t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}