 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
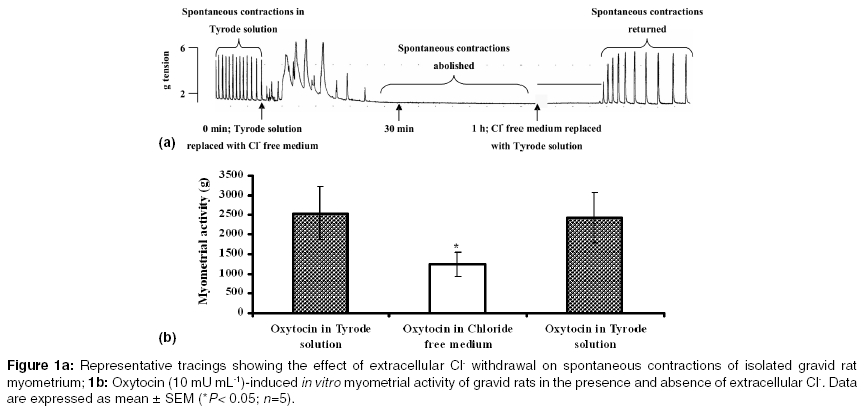
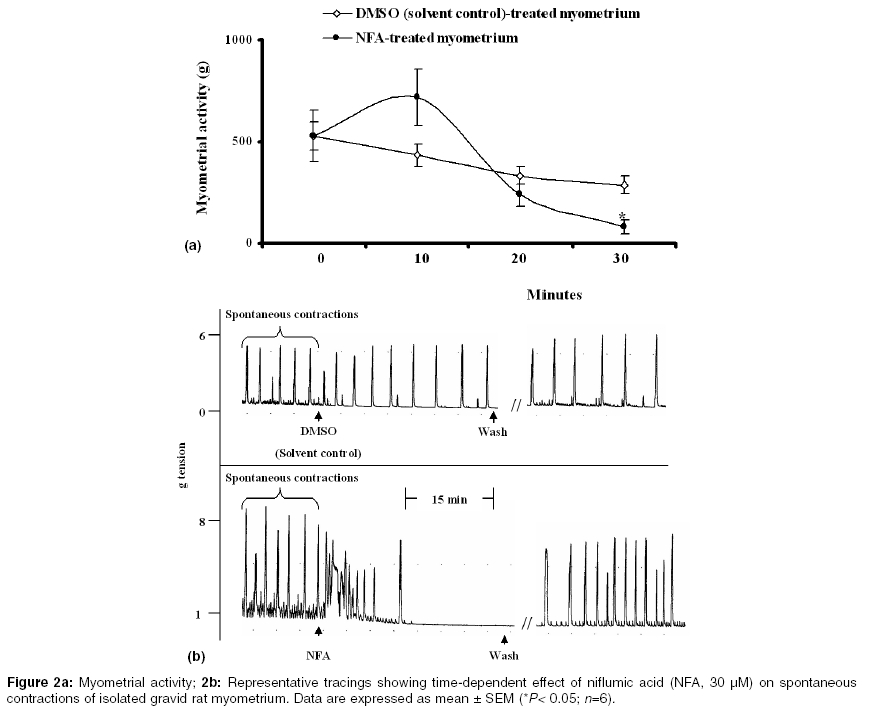
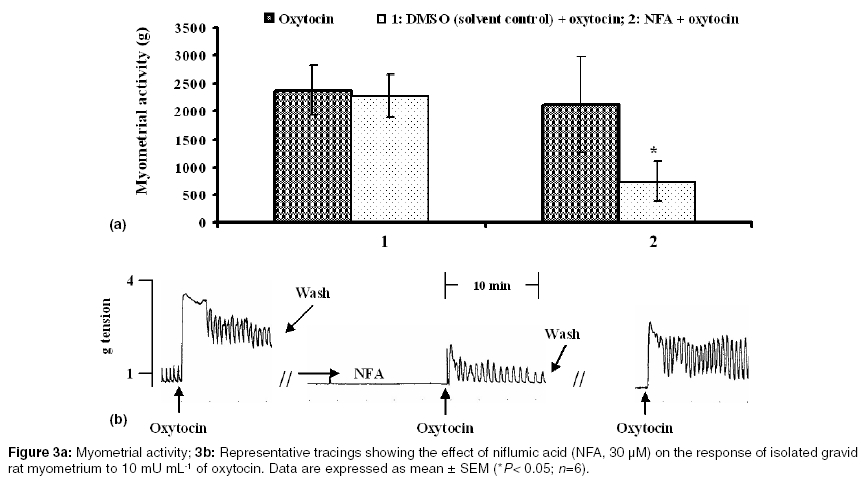
Indian Journal of Pharmacology, Vol. 37, No. 1, January-February, 2005, pp. 21-25 Research Paper Effect of functional modulation of Ca2+-activated Cl- currents on gravid rat myometrial activity Adaikan PGanesan, Adebiyi A Departments of Obstetrics and Gynaecology, National University of Singapore, National University Hospital, 5 Lower Kent Ridge Road, 119074 Correspondence Address:Departments of Obstetrics and Gynaecology, National University of Singapore, National University Hospital, 5 Lower Kent Ridge Road, 119074 obgadaik@nus.edu.sg Code Number: ph05005 Abstract OBJECTIVE: This study was carried out to examine the effect of functional modification of Ca2+-activated Cl- currents on contractile activities of isolated gravid rat myometrium.MATERIALS AND METHODS: Spontaneous rhythmic and oxytocin-induced contractile activities of gravid rat myometrium in the presence and absence of Cl- (in the bathing medium) were studied in a tissue bath system. The effects of niflumic acid (a Cl- channel blocker) on spontaneous and oxytocin-induced contractile activities were also delineated. RESULTS: Replacement of Cl--containing Tyrode solution with Cl--free medium caused transient increase in myometrial contractile tone for about 15 min (possibly due to rapid efflux of Cl-) after which gradual failure of the spontaneous contractions led to myometrial quiescence within 1 h. Normal spontaneous contractions returned after replacing Cl--free medium with Tyrode solution. In Cl--free medium, oxytocin-induced contractions of the myometrium were significantly reduced by 44 ± 9%. Niflumic acid (30 µM) caused an increase in myometrial contractile tone for about 10 min followed by time-dependent inhibition of spontaneous contractions of the rat myometrium. Moreover, prior addition of 30 µM of niflumic acid to the tissue baths (for 30 min) caused 74 ± 5% reduction of oxytocin-induced contractions of the myometrial strips. CONCLUSION: Ca2+-activated Cl- currents play a significant role in the spontaneous and oxytocin-induced myometrial activity. Ca2+-activated Cl- conductance could hence be a potential target for the pharmacological control of uterine activity. Key Words: Myometrium, niflumic acid, oxytocin Introduction It is well established that in smooth muscles, chloride (Cl-) currents are provoked by an increase in the intracellular Ca2+ concentration, leading to membrane depolarization and opening of voltage-dependent Ca2+ channels (VDCCs), which triggers Ca2+ influx and muscle contraction.[1] Calcium-activated Cl- currents (Icl(ca)) plays a role in agonist-induced contraction of vascular[2],[3] and non-vascular smooth muscles[4] and may also participate in spontaneous contractions of smooth muscles.[5],[6] Control of myometrial contractions is essential for the maintenance of pregnancy and for parturition.[7] Uterine hyperactivity before term predisposes to preterm labor - an important cause of perinatal mortality. On the other hand, inadequate uterine contractions could impede labor and increase the incidence of emergency caesarean sections.[8] Oxytocin is an important regulator of pregnancy and parturition.[9],[10],[11] Gravid rat myometrium contains calcium-activated Cl- channels.[12],[13] Using electrophysiological techniques, oxytocin was shown to activate Icl(ca) in gravid rat myometrial cells.[13] Chloride channel blockers, 5-nitro-2-(3-phenylpropylamino) benzoic acid and anthracene-9-carboxylic acid have also been demonstrated to inhibit spontaneous contractions of gravid rat myometrium.[5] It thus appears that Ca2+-activated Cl- conductance participates in myometrial contractions and could provide a potential target for the control of uterine activity. To further characterize the functional role of Icl(ca) in the regulation of uterine contractions, We studied the effect of extracellular Cl- withdrawal (from tissue-bathing medium) on spontaneous and oxytocin-induced contractions of myometrial tissue isolated from gravid, Sprague-Dawley rats. The effects of niflumic acid (a Cl-channel blocker) on spontaneous and oxytocin-induced contractions of the rat myometrium were also examined. Materials and Methods Animals The myometrial tissues employed for this study were isolated from pregnant Sprague-Dawley rats (18-20 days of gestation) housed in the Animal Holding Unit of the Faculty of Medicine, National University of Singapore. This study was conducted in accordance with National University of Singapore Guidelines and Ethics on Animal Experimentation. Chemicals and solution Oxytocin was purchased from American Pharmaceutical Partners Inc. (Los Angeles CA, USA) and niflumic acid from Sigma-Aldrich Chemical Company (St Louis MO, USA). Dimethyl sulfoxide (DMSO) and other chemicals used in the preparation of the bathing medium were purchased from Merck KGaA Darmstadt, Germany. Composition of Tyrode solution (mM): NaCl 136.9, Glucose 5.6, NaHCO3 11.9, KCl 2.68, MgSO4 1.05, CaCl2 1.8, NaH2PO4 0.37. Myometrial tissue preparations On Day 18-20 of gestation, the rats were euthanized by CO2 asphyxiation. Gravid uteri were excised to remove the fetuses and other adherent tissues and then cut into myometrial strips in a longitudinal direction. The strips (approximately 10 x 2.5 x 2.5 mm each) were mounted vertically under resting tension of 1 g weight in 4-channel (10 mL) tissue baths containing Tyrode solution. The tissue baths were maintained at 37o C, pH of 7.4, gassed with 95% oxygen, 5% carbondioxide and connected to force transducers (Ugo Basile, Comerio VA, Italy). Effect of extracellular Cl- removal on myometrial activity The myometrial tissues were washed 4 times and left to equilibrate in the Tyrode solution for at least 45 min. Following the equilibration period, the bathing medium was changed to Cl--free medium and the tissues were washed 5 times. The Cl--free medium had the same composition as the Tyrode solution, except that all the chloride salts were replaced by acetate salts. Equilibration of the myometrial strips in the Cl--free medium continued for 1 h after which the Cl- free medium was changed to normal Tyrode solution and washed 5 times. The spontaneous myometrial activity in the presence and absence of Cl- in the bathing medium was observed. The effect of oxytocin on rat myometrial tissues in the presence and absence of Cl- was also examined. This was done by contracting gravid rat myometrial strips (suspended in Tyrode solution) with 10 mU mL-1 of oxytocin for 10 min after which the tissues were washed and allowed to equilibrate in Cl--free medium for 1 h (spontaneous contractions were completely abolished at this stage). After 1 h of equilibration in Cl--free medium, oxytocin (10 mU mL-1) was added to the tissue baths for 10 min. Subsequently, the Cl- free medium was changed to normal Tyrode solution and washed 5 times. Following the rest period of at least 20 min, oxytocin (10 mU mL-1) was added to the tissue bath in order to check if the effect of extracellular Cl- removal on oxytocin-induced myometrial activity was reversible. Effect of niflumic acid on myometrial activity After equilibrating gravid rat myometrial strips in Tyrode solution, oxytocin (10 mU mL-1)-induced contractions of the myometrium were recorded for 10 min. Following the rest period of at least 20 min after wash, 30 µM of niflumic acid (NFA) was added to the tissue baths and allowed to act for 30 min (control strips were treated with equivalent volume of DMSO used in the preparation of NFA). Oxytocin (10 mU mL-1) was then added to the tissue baths for 10 min (in the presence of NFA). Thereafter, the tissues were washed and observed for recovery. The effect of NFA (30 µM) on spontaneous rhythmic contractions of the myometrium was also observed for a period of 30 min. Data analyses All responses were recorded isometrically and analyzed on MacLab computerized data acquisition system running chart v3.6 software (ADInstruments, Castle Hill, NSW Australia). Myometrial activity denotes the integral of contractions (representing tone, amplitude and frequency of contractions) during the period of contact with test substances (computed by MacLab data acquisition system). Data were expressed as the mean ± SEM. Where appropriate, Student′s ′t′ test was applied and P values less than 0.05 were considered significant. Results Gravid rat myometrium showed spontaneous rhythmic contractions few minutes after suspension in Tyrode solution. Replacement of normal Tyrode solution with Cl--free medium caused transient increase in the contractile tone for about 15 min after which gradual failure of the spontaneous contractions led to myometrial quiescence within 1 h. At about 15 min after substituting Cl--free medium with Cl--containing Tyrode solution, normal spontaneous myometrial contractions returned [Figure - 1]a. In Tyrode solution, 10 mU mL-1 of oxytocin provoked strong contractions of the myometrium. At the same concentration, oxytocin-induced activity of the gravid rat myometrium was significantly reduced (P = 0.036) in Cl--free medium (oxytocin-induced myometrial activity (g) in Tyrode solution versus activity in Cl--free medium: 2542 ± 679 versus 1232 ± 316). Subsequent to the replacement of Cl--free medium with Cl--containing Tyrode solution, oxytocin-induced contractions of the myometrium returned to 95 ± 5% of the initial activity [Figure - 1]b. NFA (30 µM) caused an increase in the contractile tone for about 10 min followed by time-dependent inhibition of spontaneous contractions of the rat myometrium [Figure - 2]. In contrast to the solvent control (DMSO), prior addition of 30 µM of NFA to the tissue baths (for 30 min) significantly reduced (P = 0.039) oxytocin-induced contractions of the gravid rat myometrium (control versus treated: 2126 ± 857 versus 729 ± 366; [Figure - 3]. Following a rest period of about 30 min after wash, the recovery of oxytocin-induced myometrial contractions was 84 ± 30% of the initial activity. Discussion One of the most probable mechanisms to provoke depolarization in smooth muscle is efflux of Cl-.[14] Activation of Icl(ca) depolarizes the smooth muscle cell to open VDCCs and the subsequent influx of Ca2+ produces contractions.[1],[6] Decades ago, Hutter and Noble[15] reported that replacement of extracellular Cl- with less-permeant anions caused transient increase in the spontaneous activity of the Purkinje fiber followed by 40-90% decrease. Similarly, replacement of Cl- with acetate (about 0.5 times less permeable than Cl-) was shown to cause efflux of Cl- and transient depolarization of sino-atrial node cells of rabbits.[16] Substituting Cl- with a less permeant anion will cause a rise in the outward electrochemical gradient for Cl- and as a result potentiate contraction.[14] As shown in this study, replacement of Cl- in Tyrode solution, with acetate caused immediate increase in spontaneous contractile tone of isolated gravid rat myometrium for about 15 min. This could be due to the rapid efflux of intracellular Cl- and depolarization as a consequence. Cl- efflux-induced increase in spontaneous contractile tone of isolated gravid rat myometrium observed in the present study was followed by total quiescence of the myometrium which was reversible after changing the Cl--free medium to Cl--containing Tyrode solution suggesting that the intracellular pool of Cl- is probably depleted (leading to failure of membrane depolarization) during the period of myometrial quiescence. Moreover, after incubating rat myometrial strips in Cl--free medium for 1 h, oxytocin-induced myometrial activity was significantly reduced. Subsequent to the replacement of Cl--free medium with Cl--containing Tyrode solution, oxytocin-induced activity of the myometrium was significantly returned to the level of its initial activity. These observations indicate that Icl(ca) play a significant role in spontaneous and oxytocin-induced gravid rat myometrial activity. The results of this functional study support the electrophysiological studies by Arnaudeau et al[13] that demonstrated oxytocin-induced chloride currents in pregnant rat myometrial cells. NFA, a rapidly dissociating open channel blocker[17] has been suggested to be one of the most potent inhibitors of Icl(ca).[6] In the present study, incubation of rat myometrial strips in 30 µM of NFA for 30 min caused significant but reversible inhibition of oxytocin-induced activity of the myometrium. This inhibition occurred at a concentration within the range of those earlier shown to selectively inhibit Icl(ca).[4],[18] However, NFA (30 µM) caused transient increase in the tone of spontaneous rhythmic contractions of the myometrium within 10 min of addition to the tissue baths. After about 10 min of incubation in NFA, spontaneous activity of the rat myometrium was inhibited in a time-dependent fashion. NFA-induced increase in the tone of spontaneous contractions within 10 min of its addition to the tissue bath was unanticipated of a Cl- channel blocker. Nonetheless, using rabbit pulmonary artery smooth muscle cells, Piper and colleagues[19] recently demonstrated for the first time that NFA has dual effects on Icl(ca). The mechanism of the dual effect of NFA on Icl(ca) is not completely understood, however, Piper et al[19] suggested that NFA increases Icl(ca) and at the same time inhibits Cl- conductance by a channel blocking mechanism and the net result is dependent on the direction of ion flux. It is thus possible that the increase in the spontaneous activity of the myometrium observed within 10 min of addition of NFA to the tissue baths was caused by NFA-enhanced Icl(ca), while the inhibition of the spontaneous activity of the myometrium after 10 min of addition of NFA was due to Icl(ca) blockade. Further investigations are necessary to prove this supposition. In conclusion, Ca2+-activated Cl- currents appear to play a significant functional role in spontaneous and oxytocin-induced myometrial activity. Modulation of Ca2+-activated Cl- conductance could hence provide a potential target for pharmacological control of uterine activity. References
Copyright 2005 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph05005f2.jpg] [ph05005f3.jpg] [ph05005f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}